


Abstract
Grapevines are grown either on their own roots or on rootstocks that represent a mixture of grapevine species and hybrids. Developmental and physiological factors other than phylloxera resistance, lime tolerance, and ease of propagation were not directly considered during rootstock breeding, including rooting patterns. Here, in a comprehensive literature synthesis, we have compiled information concerning rooting depth distributions of grapevine roots from wall profile studies comprising a broad range of soil environments and rootstock genotypes. We considered the distributions based on the asymptotic equation of Y = (1 − ßd), where d = soil depth (cm) and Y = the proportion of roots from the surface to depth d. The median value of ß for the root distributions analyzed was 0.9826 and the standard deviation over all observations was 0.0068 (n = 240); most profiles had fitted values of ß generally greater than 0.975. This value places the depth distribution of grapevine roots in the vadose zone among the deepest observed for plants worldwide. The data suggested that soil properties such as the presence of soil profiles impermeable to root penetration, stoniness, and presence of gravel lenses have a greater influence on depth distributions than does genotype, even in deep fertile soils. Genotypic differences were not apparent, although the rootstock O39-16 (ß = 0.9867 ± 0.0009, mean ± se, n = 11), with a reputation for deep-rooting behavior, did exhibit deeper root distributions. The analysis also suggests that root characteristics other than root horizontal and vertical spread may need to be considered in order to explain some key rootstock characteristics like scion vigor or drought tolerance.
Roots provide structural support and surface area for water and mineral nutrient absorption. For grapevines and many other lianas, the existence of tendrils (Mullins et al. 1992) curtails the need for roots as structural support organs. Consequently, the evolution of grapevine root form and function has probably been directed by factors other than support, like demands for water and nutrient foraging. For the rootstock species and hybrids within the genus Vitis, the size of the root system, in terms of its horizontal and vertical depth distribution, is important to water and nutrient foraging capacity. Variation in vertical and horizontal depth distribution of grapevine root systems may have a genetic component (Guillon 1905, Pongrácz 1983) (Figure 1⇓), while environmental parameters controlling phenotypic expression may play a more definitive role (Perold 1926, Winkler et al. 1974, Van Zyl 1988). Teasing apart the relative contributions of environmental versus genetic controls on root distributions of grapevines and then understanding functional differences among the diverse genotypes that represent grapevine rootstocks will be extremely challenging.

Emergence angles of adventitious roots produced by cane cuttings of, from left to right, Riparia Gloire de Montpelier (V. riparia Michaux), V. riparia x V. rupestris cv. 3309C, and Rupestris du Lot (V. rupestris Scheele) (Guillon 1905; reproduced with the permission of Masson Publishers/Dunod, Paris).
Work conducted primarily in South Africa has suggested that variation in vine form and function aboveground (mainly size) may serve as an indicator of characteristics like soil water-holding capacity or fertility below ground (Saayman and Van Huyssteen 1980, 1983, Van Zyl and Van Huyssteen 1980, Archer and Strauss 1985, Archer et al. 1988, Van Huyssteen 1988a, Morano 1995). Distinct and sometimes predictable growth patterns are known to emerge for specific rootstock/scion combinations, and the phenotypic patterns expressed are thought to contribute to the physiological performance of the whole vine (Pouget 1987). These “truths” seem to be widely accepted among viticulturists, but direct experimental verification is generally lacking. Evidence from nonagricultural systems may support this hypothesis, where lateral root spread and maximum rooting depth are significantly correlated with aboveground plant volume (Schenk and Jackson 2002). Nonetheless, in this case (Schenk and Jackson 2002) it was evident that plant growth form—grass, herb, shrub, tree—was the primary factor controlling rooting depth, even though strong correlations emerged between rooting depth and mean annual precipitation. Other comprehensive reviews of rooting depth with a focus on woody perennials have emphasized the role of soil properties (root impenetrable layers, anoxia, and the existence of shallow soils over bedrock) rather than plant form in controlling rooting depth (Stone and Kalisz 1991).
The report of Seguin (1972) is one of the most widely referenced articles concerning grapevine root depth distribution (Richards 1983, Mullins et al. 1992), but consists of the single observation of a root growing to a depth of 6 m in the Bordeaux region of France. Other reports exist where root lengths >6 m have been documented (Doll 1954, Branas and Vergnes 1957), so it is evident in some soils that grapevines can have very deep maximum rooting depths and depth distributions. It is likely a genotypic component contributes to this as grapevine canes can extend over time to reach lengths >25 m (A. Lakso, unpublished data, 2004). As numerous other woody plant species have been shown to have roots that extend to lengths as great as 60 m (Stone and Kalisz 1991, Canadell et al. 1996, Schenk and Jackson 2002), it is self-evident that grapevine roots will eventually be reported at greater depths than those reported by Seguin (1972). How maximum rooting depth relates to the proportions of grapevine roots distributed horizontally and vertically in the upper one to two meters of soil has not been addressed. Many reports of grapevine root distribution patterns exist (see Appendix⇓), but few observations of maximum rooting depths have been reported. Historically, the goals of investigations of depth distributions have been to document differences among rootstock genotypes in a particular soil type or to document the influence of various management practices on rooting patterns. These reports have never been reviewed, with the exception of a compilation of South African research (Van Zyl 1988). The purpose of this article is to comprehensively review the published data on vertical and horizontal root distributions of the species and hybrids of Vitis growing in diverse soil environments.
Appendix: Root Distribution Table
Methods for Studying Grape Root Distributions
There are several methods of studying root distributions, but the most commonly accepted in viticulture has been the profile wall method (Bohm 1979). As observed by Jackson et al. (1996), the earliest recorded profile wall studies of crop plant roots were attributed to Schubart (1857), who examined hydraulic excavations of crop roots from wall profiles, and to Sachs (1873), who used a glass rhizotron wall. The earliest report to our knowledge of grape root distributions was more recent (Harmon and Snyder 1934), but the authors employed a large-scale excavation. The actual use of profile walls to study grapevine roots did not first appear until much later (Branas and Vergnes 1957, Howarth 1959), and with the previously unpublished work of Nelson Shaulis, who examined an extensive number of profile wall distributions from 1960 to 1963. To our knowledge the first profile wall distributions used to study grapevine roots following those of Shaulis did not appear until the early 1980s in studies conducted in South Africa (see Van Zyl 1988 for a review).
The profile wall method typically consists of excavating a trench of 1- to 2-m depth at some predetermined location generally parallel to the vine row, establishing a grid of fixed subquadrat areas on a wall of the trench, and then recording root-wall intercepts. In order to improve visibility, the plane of the wall is sometimes painted white and the soil subsequently teased away from roots emerging from the wall (Van Zyl 1988). In most cases, the profile wall is established approximately 0.3 to 1 m from the vine trunk, although there seems to be no preferred distance. Morlat and Jacquet (1993, 2003) have promoted the use of wall profiles excavated at sequentially decreasing distances from the trunk, but few additional reports exist where researchers have adopted a similar approach (but see McKenry 1984). A profile can also be established on the floor of successively deepened trenches (Morlat and Jacquet 1993), but again the use of multiple layered excavations are rare in comparison to single wall profiles at a set distance from the trunk.
There are several shortcomings to the wall profile method. One is the explicit assumption that the wall profile being examined bears relation to root intercepts that would be observed if the plane were measured at the same distance from the trunk, but set in another compass direction other than the most common, which is parallel to the vine rows. In other words, there is an assumption that the 3-dimensional distribution of roots around the vine is relatively uniform. This assumption is probably not always true. Nonetheless, there are several reports that support such a contention. Kozma (1967) did extensive drawings of excavations apparently attributed to mapping of rootstock 110R that suggest a uniform spread of roots (Figure 2⇓). In addition, at least three investigations used wall profiles set in both a parallel and perpendicular direction to the trunk, but at the same distance from the trunk (Williams and Smith 1991, Padgett-Johnson 1999, Morlat and Jacquet 2003). Most of these investigations reported similar depth distributions in both the parallel and the perpendicular planes (Appendix⇑).

(A) Vertical root distributions for V. berlandieri x V. rupestris cv. 110R from the work of Kozma (1967). (B) Horizontal root distributions of V. berlandieri x V. rupestris cv. 110R from the work of Kozma (1967).
One interesting exception to the observation of similar distributions in parallel and perpendicular excavations was observed in a study by Padgett-Johnson (1999) of a wild accession of V. champinii. It had over 95% of roots in the upper 60 cm of soil in a 1-m square area within the berm of the vine row and trenched perpendicular to the vine row. This percentage was unusually high when compared with all other root wall profiles we examined and may suggest that V. champinii roots cluster in the drip-zone area even when not growing in shallow, dry soils. Nonetheless, the study consisted of a single observation. Further and more extensive observations on V. champinii accessions or hybrids are needed to support this hypothesis. If verified, then V. champinii may constitute a good genotype for examining relations between emergence angles and depth distribution. Despite this exception we found excavations conducted in relatively deep, fertile, and uniform soils without substantial spatial discrepancies in terms of the existence of stones, gravel lenses, or clay profiles yielded relatively uniform root distributions overall (see Figure 3A⇓ and Appendix⇑).
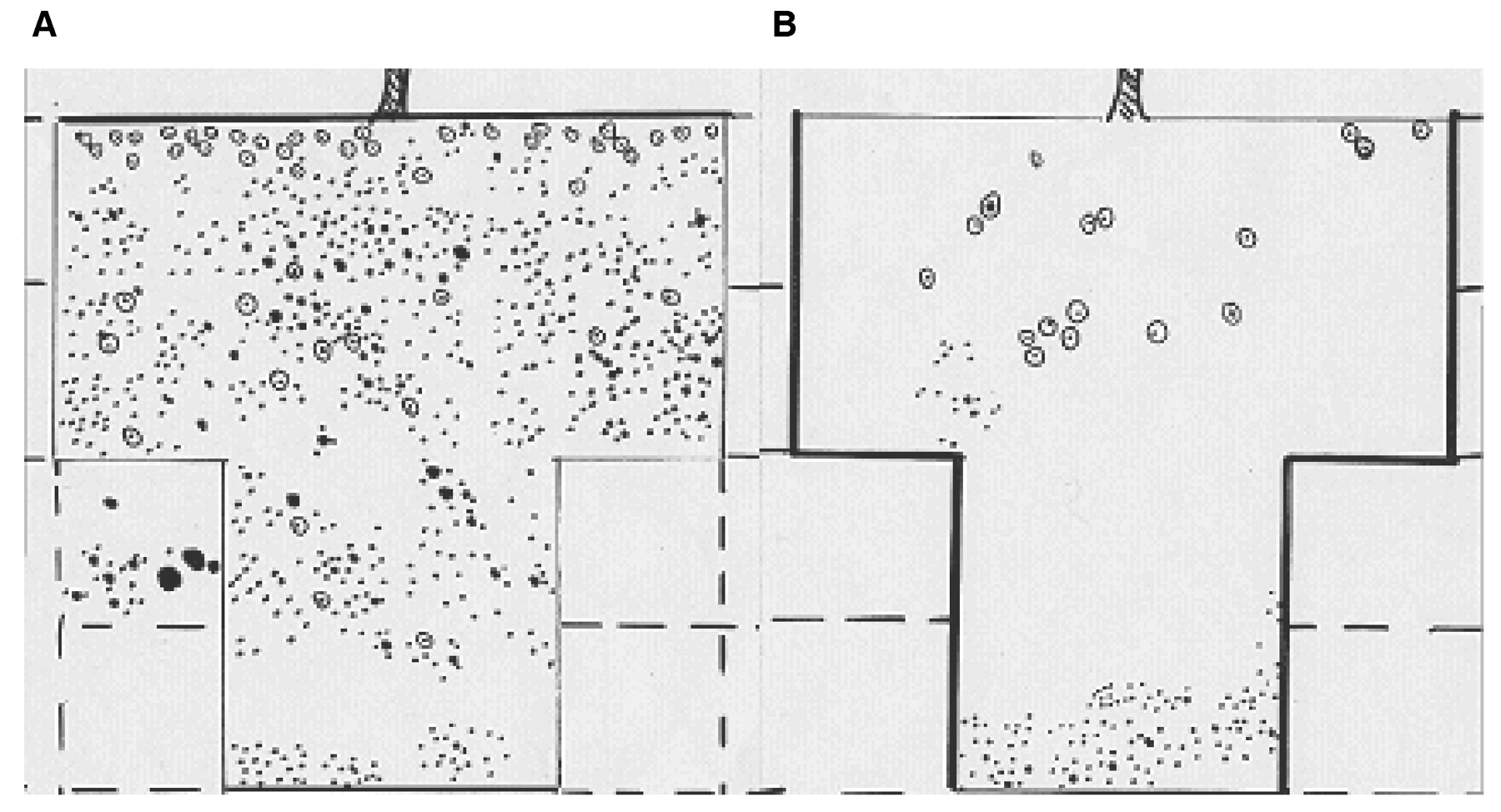
Root wall profiles from (A) a Chenango sandy clay loam soil in the Finger Lakes region of New York and (B) a Chenango stony sandy clay loam soil with a gravel lense in the lowest profile level (N. Shaulis, unpublished maps, 1963; reproduced with permission of the New York State Agricultural Experiment Station).
Another shortcoming to wall profiles concerns the observation that penetration of a barrier by a few roots can lead to substantial root proliferation below that zone (Figure 3B⇑), given adequate aeration and moisture (Kimber 1974, Armson 1977). A similar kind of nonuniformity can occur when roots encounter areas where mechanical resistance to root elongation is diminished and water and nutrients are available. On a more practical level, the possibility that substantial heterogeneity exists in 3-dimensional distributions (Figure 3A, B⇑) may be less important than the primary information sought by viticulturists, the main one being maximum depth to which roots can grow.
Finally, a profile wall may not reflect the viewing of roots of a single or a pair of target vines in the vineyard (McKenry 1984, Morano 1995). Since the horizontal and vertical spreads of grapevine roots can reach distances of 3 m to >8 m (Doll 1954, Branas and Vergnes 1957, Kozma 1967) (D. Smart, unpublished data, 2002), there is likely a tremendous overlap of roots in vineyards, even those with relatively wide plant and row spacing (see Figure 2B⇑). It is also likely that interspecific root-grafting (Stone and Kalisz 1991) occurs among individual vines within vineyards. For these reasons, we believe a vineyard below ground in many respects may be considered a community of roots of clonal individuals. If extreme nonuniformity is not apparent in the horizontal spread of grapevine roots (Figure 2B⇑), then the position of the viewing plane with respect to an individual vine should not strongly influence results concerning depth distribution. This point of view is supported by the detailed excavations performed by Horvath (1959) showing strong overlap of roots among neighboring vines (Figure 2B⇑). To the contrary, Archer and colleagues (Archer and Strauss 1985, Archer et al. 1988) found that an increased planting density increased penetration angle of roots emerging from the trunk. They also reported that an increased planting density diminished root-system size while increasing root density (Archer and Strauss 1985). Morano (1995) reported increased rooting depth and increased root density (number per wall area) as vine spacing decreased from 2 to 1 m, and thus maintained that closer vine spacing resulted in a smaller, denser root system. We conclude that more information is needed on how plant density influences the size and horizontal and vertical spread of grapevine roots. In addition, further information is needed concerning how heterogeneous environments in terms of resource availability, mechanical resistances, and vine density influence the size and distribution of roots.
Methods
We conducted an extensive search for reports of root distributions for Vitis species and hybrids and closely related taxa such as Muscadinia rotundifolia. These reports included journal articles, book chapters, technical reports, and unpublished data contributed by colleagues in the form of notes and memoirs, particularly those of Nelson Shaulis. Although we tried to be as thorough as possible, it must be noted that such information is often published in trade journals, special publications, and proceedings, which can be difficult to fully access through standard scientific indexing services. In addition, it was challenging to search and access older information, especially that originating from outside North America. Information on root distributions for grapevines is often reported as ancillary information to projects with goals other than rooting behavior. As a consequence, more information than we have included in this report concerning grapevine root distributions may exist. Nonetheless, we judged that the data assembled in the Appendix⇑ is great enough to provide extensive and useful information and to address several assumptions about grapevine rooting behavior, even though it may not be complete.
Data were used for this analysis if roots were divided and quantified in at least four separate horizons and to a minimum depth of 75 cm. There were few cases where this was not possible, and in some cases maps were available where the soil profile descriptions had also been included. We divided the mapped regions into subquadrat areas of 10 cm depth and quantified root distributions where counts did not exist in the published or unpublished studies. The cumulative fraction of roots with increasing depth was calculated. The cumulative fraction was then expressed as a function of soil depth by testing its fit to the model Y = (1 − ßd) where Y is the cumulative fraction of roots with depth and d is the soil depth in centimeters. The coefficient ß can be used as a numerical index of depth distribution where higher values for ß correspond with greater proportions of roots with depth (Gale and Grigal 1987, Jackson et al. 1996).
The data from all the references were organized into an appendix that would allow us to examine it according to genotype, soil type, and soil properties, as well as climatic region and variables such as annual rainfall. The data on soil type, for example, were not always easy to derive because it was often difficult to establish a precise location where the excavation was made. Soil properties and even soil type can easily change within a vineyard, and for this reason only generalized information was available for each excavation or for each excavation set.
Results and Discussion
Vertical distribution of grapevine roots.
We encountered over 200 trench-wall profiles from approximately 40 different species and hybrids of Vitis and Muscadinia reported during the previous 40 years (Appendix⇑). Nearly all of the wall profiles we analyzed yielded statistically significant fits (p < 0.05) to the equation describing an exponential rise in root fraction with soil depth, Y = (1 − ßd). The exceptions occurred for one own-rooted Vitis labruscana cv. Concord vine and a 3309C rootstock in eastern North America around the Finger Lakes region. In these two cases, significant fits were obtained using an exponential model Y = ßd. Nevertheless, there were other data sets where exponential models fit the data as well as the model Y = (1 − ßd). These distributions did not appear to reflect genotypic or climatic differences. Rather, where soils contained large stones, clay layers, gravel lenses, or other soil profile changes either impermeable to roots or, conversely, highly permeable to roots, root distributions became patchy with greater root densities occurring in defined areas at depth (Figure 3B⇑). Nearly all of the root profiles we encountered were taken from relatively deep fertile soils, and in such cases root distributions were clearly more uniform than from stony or gravelly soils.
Most of the values for ß we observed exceeded ß = 0.976, the average value obtained by Jackson (1996) for temperate coniferous forests, the biome they reported to have the deepest root distributions. The average value of ß over all species and hybrids of grapevine we analyzed was 0.9826, with a standard deviation of 0.0068 (n = 240). The fits to the model indicated that approximately 63.2 ± 2.6% (mean ± 95% CI, n = 240) of grapevine roots were in the upper 60 cm, and 79.6 ± 2.4% (mean ± 95% CI, n = 240) within the upper 1.0 m. For the coniferous forests (Jackson et al. 1996), the percentage of roots encountered to the same depths was 76.7% and 91.2%, respectively. Thus, grapevines as a group appeared to have proportionally deeper root distributions in the vadose zone compared with many plants in natural ecosystems. It must be kept in mind that grapevine root distributions reported are from selected, disturbed, and managed agricultural systems. Factors such as cultivation or altered competitive relationships may influence the above results compared with undisturbed natural ecosystems. This observation also does not indicate that grapevines have extremely deep roots per se. Many other plants have been reported to have roots up to 50 meters longer than the longest reported for grapevine (Stone and Kalisz 1991, Canadell et al. 1996). The maximum rooting depth of grapevines and how maximum rooting depth relates to vine performance are yet to be determined. Given the depth of grapevine roots in the upper one to two meters of soil, we expect their roots to reach depths comparable to those reported for other woody taxa.
Lateral spread of grapevine roots.
Investigations characterizing lateral spread of grapevine roots were rare. Studies that did so generally relied on arrays of soil cores from which root length densities or fresh weights of roots were recorded. In at least three cases, detailed excavations were undertaken (Horvath 1959, Kozma 1967, Saayman and Van Huyssteen 1980, McKenry 1984, Morlat and Jacquet 2003). These researchers generally found fairly high root densities at distances greater than a meter from the vine trunk. Although Saayman and Van Huyssteen (1980) found lateral spread was somewhat restricted in a soil previously ripped along the vine row, they nevertheless found that root densities were still relatively high at 1.5 m from the trunk.
Nagarajah (1987) quantified root length densities into the row for own-rooted V. vinifera cv. Thompson Seedless and Thompson Seedless grafted onto V. champinii cv. Ramsey rootstock. He used an array of cores sampled at 30, 90, and 120 cm into the vine row from the trunk, and in 10-cm increments to 120 or 220 cm depth, depending on soil texture. He found root length densities to range between 0.4 and 1.7 mm cm− depending on soil texture, with coarse-textured soils having the lowest root densities and fine-textured soils having the highest densities. Horizontal root length densities suggested that the spread of these two genotypes was fairly extensive with respect to the areas sampled, with density diminishing by an average of only 28.0 ± 5.4% (mean ± se, n = 9) from 30 to 90 cm distance from the trunk. Perry et al. (1983) used a slurry method to quantify root length densities in samples extracted from cores taken in the row for four grape species (Appendix⇑). They found unacceptable variation in core samples as compared with wall profiles and reported the technique to be unreliable. McKenry (1984) examined lateral spread of roots in a Thompson Seedless vineyard using root length and fresh mass. He found that approximately 11.6% and 14.4% of the total root biomass were found 1.2 to 1.5 meters from the trunk when excavated to a depth of 1.2 m with respect to a raised berm (Figure 4⇓). This was actually a fairly large proportion considering roots excavated near the trunk were large framework roots and thus would weigh more. The detailed drawings of Kozma (1967) indicated that some larger framework roots can reach a maximum spread of approximately 10 m, and our own excavations support this contention (D. Smart, unpublished data, 2002).
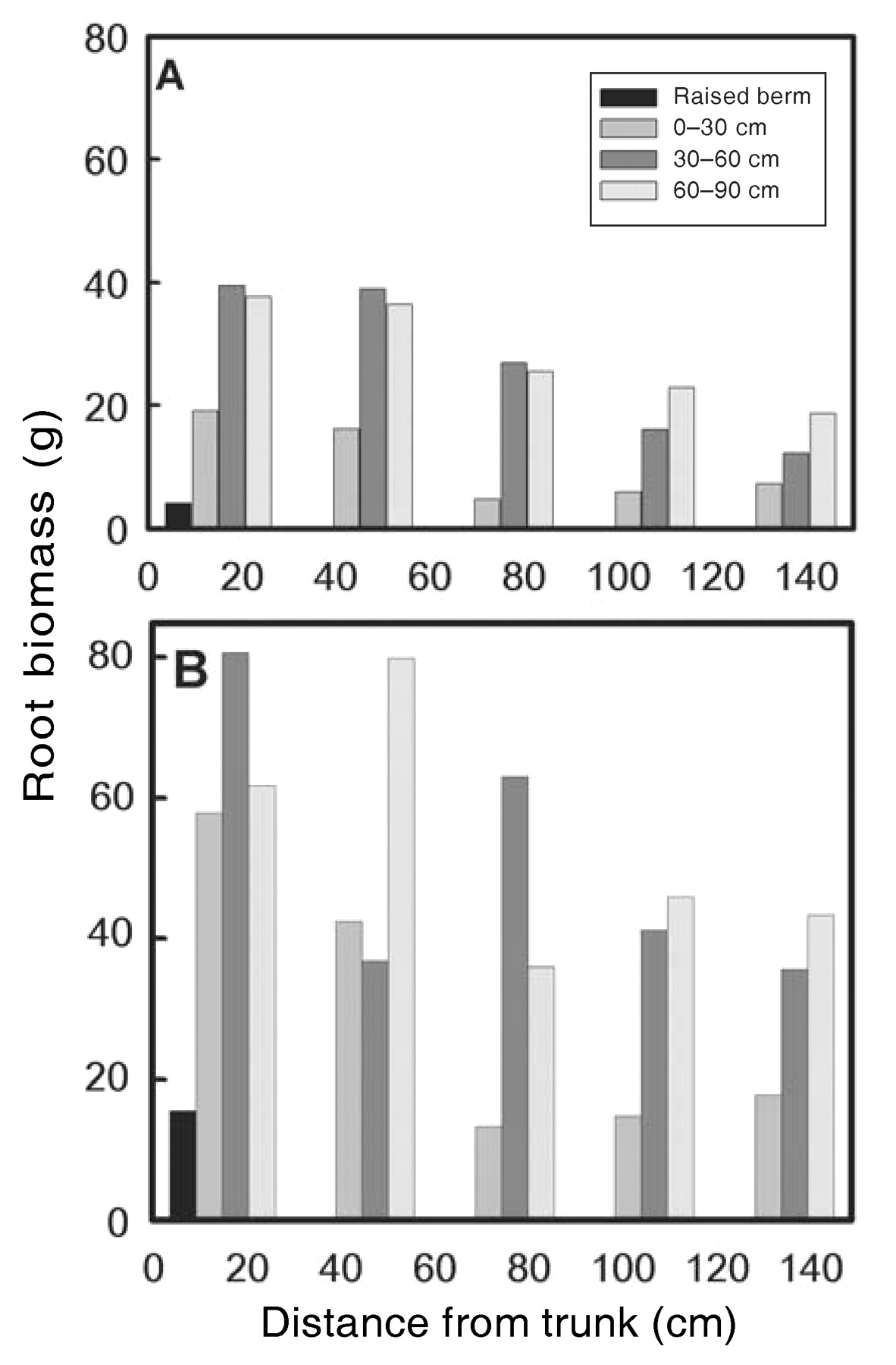
Root fresh biomass (g m2) for (A) V. vinifera cv. Thompson Seedless and (B) V. solonis X V. rupestris cv. Ramsey (McKenry 1984).
Soil physical properties and grapevine roots.
Soil texture may influence rooting patterns in the sense that fine-textured soils would have higher water-holding capacities, lower resistances to water extraction, and shallower infiltration rates than coarse-textured soils (Brady and Weil 2002). So, it might be predicted that root systems in fine-textured soils would be smaller and shallower and, conversely, those in coarse-textured soils deeper (Sperry et al. 1998, Jackson et al. 2000). Even so, correlations between texture and horizontal or vertical spread in natural ecosystems have not been found (Schenk and Jackson 2002).
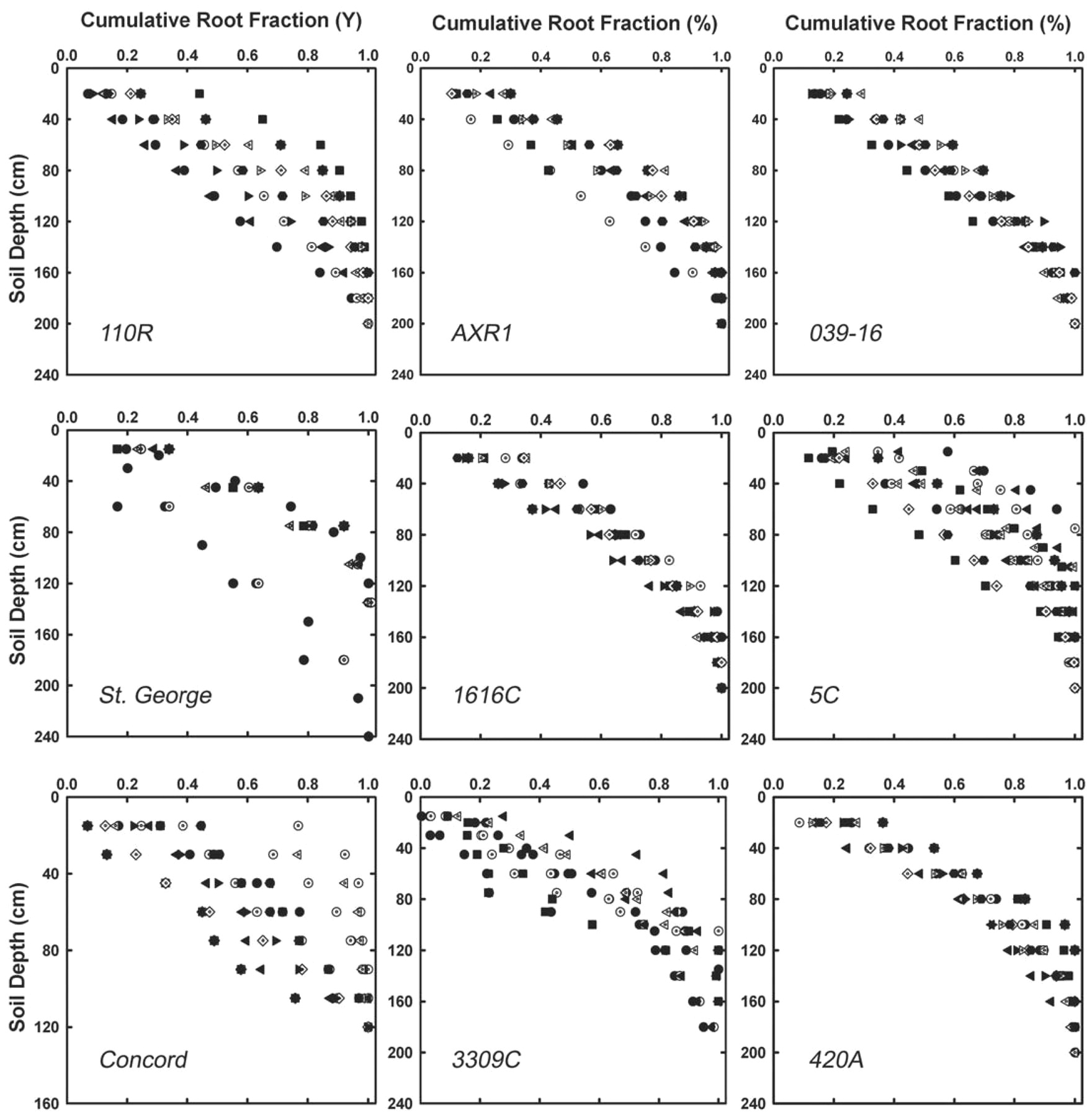
The data examined in this review (Appendix⇑) also did not fully support that texture strongly influenced rooting depth distributions, although Nagarajah (1987) observed that root proliferation by Thompson Seedless was deeper in coarse- textured soils compared with fine-textured soils. However, Morlat and Jacquet (1993) and Araujo et al. (1995) found that root densities were simply diminished in the upper 0 to 20 cm of sandy soils, which could give the impression of deep rooting behavior. It is possible that rapid drying and extreme temperatures may shorten root lifespan in these shallow soil layers rather than that roots penetrate deeper. The root wall profiles we analyzed indicated that layers resistant to root penetration like those with high bulk densities (Saayman and Van Huyssteen 1980, Van Huyssteen 1988a) influenced depth distribution regardless of textural class. Thus, it may be that coarse-textured soils, or those that are clean cultivated (Van Huyssteen and Weber 1980), are simply devoid of roots in surface soils rather than roots penetrating to deeper soil layers. In addition, Vitis rupestris cv. DVIT 1263 growing in a deep sandy loam soil (coarse texture) in the Central Valley of California (Padgett-Johnson 1999) had a very similar root depth distribution (ß = 0.9830, n = 1) to V. rupestris cv. St. George growing in deep, clay loam soils (fine texture) in the Carneros region of Napa Valley (Sipiora et al. 2005) (ß = 0.9822 ± 0.0006, mean ± se, n = 4). Both were deeper rooted than St. George growing in a gravelly clay loam in the Oakville region of Napa Valley (Morano and Kliewer 1994) (ß = 0.9780 ± 0.0006, n = 4). Williams and Smith (1991) found St. George to be profoundly deep rooted in the same gravelly clay loam soil in Oakville (ß = 0.9918 ± 0.0022, mean ± range, n = 2), although the root densities they reported were extremely low compared with other reports. Findings regarding rooting by this genotype and textural class were therefore inconclusive (Figure 5⇓).

Cumulative root distributions (cumulative fraction of the total) as a function of soil depth for nine rootstock genotypes. The data in each panel can be described by the least squares fit of the theoretical model of Gale and Grigal (1987) where Y = (1 − ßd), where Y is cumulative fraction of roots with depth and d is the soil depth (cm) as follows: 110R = 0.9843 ± 0.0018 (mean ± se, n = 14); AXR1 = 0.9850 ± 0.0011 (n = 18); 039-16 = 0.9867 ± 0.0006 (n = 11); St. George = 0.9822 ± 0.0016 (n = 9); 1616C = 0.9835 ± 0.0011 (n = 17); 5C = 0.9832 ± 0.0014 (n = 20); Concord = 0.9813 ± 0.0052 (n = 17); 3309C = 0.9842 ± 0.0013 (n = 19); 420A = 0.9840 ± 0.0011 (n = 10).
The studies cited above, and others, illustrated that soil structure, stoniness, and depth to the water table were the key determinants of vertical root distribution regardless of genotype or texture (see Figure 5⇑ and Appendix⇑). Using St. George again as an example, in a moderately deep, clay loam soil in the Carneros region of Napa Valley, St. George (V. rupestris) had roots reaching a depth of 150 cm and ß = 0.9822 ± 0.0006 (Sipiora et al. 2005); whereas, in the deep, gravelly clay loam soil with a high seasonal water table, the rooting depth was reportedly only 120 cm and the observed value of ß was lower (0.9780 ± 0.0006). For rootstock 5C, ß was 0.9846 ± 0.0017 (n = 15) in gravelly clay loam soils in the Napa Valley, while in the Finger Lakes region in a soil that was extremely stony in the lower soil profiles, ß was 0.9609 ± 0.0069 (mean ± range, n = 2) (see Appendix⇑).
Plant available water is determined both by texture and rooting depth. It is likely a combination of these variables, in addition to climatic factors such as mean annual precipitation, might provide better insights into controls on rooting depth distribution. In viticulture science, it is generally perceived that soil depth is the more important determinant of rooting depth distribution (Van Zyl 1988), and this perspective is supported by Saayman and Van Huyssteen (1980, 1983). Grapevine roots have relatively low densities in soils (Morano 1995) and extensive lateral and vertical spreads (this review). Thus, grapevines may colonize a more extensive rooting zone at low rooting densities than do some plant functional types considered in more general analyses, such as grasses, herbs, and shrubs.
Saayman and Van Huyssteen (1983), working in a heavy Sterkspruit soil, found that high bulk densities that impede root penetration were the most important factors in limiting root downward penetration and yields. They observed that rooting depths of less than 60 cm were not amenable to developing a root system capable of sustaining optimal vine productivity under dryland conditions. Our analysis may support this contention, in that only 63.2 ± 2.6% of roots were found in the upper 60 cm when vines were growing in mostly deep, fertile soils. Grapevines may have evolved to develop deep root systems with relatively low overall densities, which may increase the probability of encountering available resources. For example, their roots may be able to reach deep or distant water sources, and then employ hydraulic redistribution to sustain the vine and provide moisture for evapotranspiration (Smart et al. 2005). This hypothesis would fit well with the point of view that grapevine root systems evolved under intense competition with their host trees (Morano 1995). Further support of this hypothesis comes from the work of McKenry (1984), who found that grapevine roots rapidly proliferate into biopores created by dead and decomposing roots or into natural fracture lines.
Influence of cultural practices on grapevine root distributions.
A number of the root wall profiles in the Appendix⇑ were taken from investigations that examined the influence of cultural practices on grapevine root distribution patterns (Van Huyssteen 1980, 1988a,Van Huyssteen b, Saayman and Van Huyssteen 1983, Morlat and Jacquet 2003). Some consistent observations on rooting patterns have emerged from those investigations. First, several studies have consistently noted that clean cultivation through tillage or the establishment of a permanent sward greatly diminishes root presence in the upper approximately 20 to 30 cm of soil (Figure 6⇓), depending on depth of tilling and cover-crop type (Saayman and Van Huyssteen 1983, Soyer et al. 1984, Ancel 1986, Morlat and Jacquet 2003). Root pruning is probably the primary mechanism eliminating roots in the upper profiles in the case of clean cultivation, but see Van Huyssteen (1988b) for a discussion of how soil physical conditions unfavorable for grapevine root growth are also strongly influenced by tillage practices. In the case of cover crops, it is generally accepted that reductions in soil moisture content and intensive competition with cover-crop roots account for the absence of grapevine roots in the upper profile. It remains to be determined whether or not this is a consequence of accelerated mortality or is due to roots avoiding growth into areas where competition from cover-crop roots would occur. If, in fact, avoidance is the mechanism driving root absence, this observation again supports the hypothesis that grapevine roots may have evolved to be effective competitors with their host trees through deep-rooting behavior.

Root distributions for V. rupestris x V. riparia cv. 101-14 Mgt under four different cultivation treatments (Van Huyssteen and Weber 1980; reproduced with permission of the South African Journal of Enology and Viticulture).
Minimum tillage practices such as the application of mulches consisting of organic material (Van Huyssteen and Weber 1980) or plastic (Van Huyssteen 1988b) or the application of herbicides can increase root densities in the upper 20 cm of soil. Soil moisture retention is thought to explain either improved root survivorship or proliferation in the upper soil profile, but diminished soil temperatures resulting in lengthened root survivorship cannot be ruled out. At least one investigation has reported on the influence of deep ploughing or ripping on root depth distributions (Saayman and Van Huyssteen 1980), but the study was inconclusive. This was partly due to the presence of a duripan at 70 to 100 cm that was unaffected by either the ripping or the ploughing treatment but that limited root distribution. A further drawback to this study, as was true for many of the reports we encountered, was the absence of sufficient repetition and therefore our ability to statistically separate means.
Influence of genotype on grapevine root distributions.
The existence of genotypic variation in rooting behavior by Vitis rootstocks has received limited attention with the exception of attempts to examine rootstock rooting patterns under nonexperimental field conditions (Southey and Archer 1988, Swanepoel and Southey 1989, Morano 1995, Padgett-Johnson 1999). Most of these studies were inconclusive and, again, were hampered by lack of repetition and/or extreme heterogeneity in root distribution of individual vines. Swanepoel and Southey (1989) did find somewhat compelling evidence that V. berlandieri cv. 13/5, V. berlandieri x V. rupestris cv. 1103P, V. riparia x V. rupestris cv. 101-14 Mgt, and 110R had greater root densities to depth than other rootstocks they examined. Our analysis indicated that all of these rootstocks had similar rooting depth distributions (see Appendix⇑), suggesting that root density rather than rooting depth per se may be a key difference among rootstocks with diverse performance in terms of scion growth.
The data we reviewed do not fit with some of the first attempts to characterize genotypic variation in grapevine rooting patterns. Guillon (1905) attempted to sort rootstock species and hybrids into those whose roots penetrated deeply (sinker roots) and those whose roots would grow more horizontally in subsurface soils (feeder roots). Guillon examined this phenomenon by measuring the geotropic response, or emergence angles, of roots growing out from the basal node of cane cuttings (Figure 1⇑). He reported that genetic differences existed in emergence angles: V. riparia cv. Riparia Gloire was reported to have the steepest emergence angles from the vertical line of the cane, 3309C had an intermediate angle, and Rupestris du Lot (St. George) exhibited more shallow emergence angles (Figure 1⇑). Our compilation of known data for rootstock distribution patterns indicates that when growing in large unrestricted soil volumes, emergence angles from stem cuttings may not be a good indicator of the depth distribution of roots (Appendix⇑). The rootstock 3309C growing near Fredonia above the Finger Lakes region was one for which we found the largest number of reported observations for rooting depth distributions. These distributions had an extremely wide range of ß values, with ß = 0.9651 ± 0.0073 (mean ± range, n = 2) in a stony, clay loam soil to ß = 0.9867 ± 0.0035 (mean ± se, n = 5) in a neighboring clay loam where texture changed with depth (clay loam at 0 to 30 cm and sandy clay loam at 30 to 90 cm).
Individual taxa within grapevine did sometimes adhere to commonly accepted ideas about grapevine root distributions, but in other cases did not. For example, we expected the rootstock V. vinifera x Muscadinia rotundifolia cv. O39-16, to be deep rooted (A. Walker, personal communication, 2004). The values of ß we observed (ß = 0.9867 ± 0.0009, mean ± se, n = 11) agreed with this observation. However, V. berlandieri x V. riparia cv. 5C, a root-stock that generally confers low vigor and performs well in soils with poor drainage, also exhibited relatively deep rooting behavior in most soils where 5C rootstock was examined (ß = 0.9832 ± 0.0014, n = 20). A shallow rooting behavior would seem more likely for the 5C genotype in terms of its ability to thrive in wet soils and its reputation for diminished vigor, an idea frequently put forth. Nonetheless, the only soil where 5C was relatively shallow rooted was a Chenango sandy clay loam where ß = 0.9609 ± 0.0069 (n = 2, mean ± range), and approximately 90% of the roots were found in the upper 60 cm (N. Shaulis, unpublished research, Cornell University, 1963). Furthermore, V. berlandieri x V. riparia cv. 420A and cv. 110R, also not known as high-vigor conferring rootstocks, had relatively deep root profiles as well, with ß = 0.9840 ± 0.0011 (mean ± se, n = 10) for 420A, and ß = 0.9843 ± 0.0017 (n = 14) for 110R. The above observations taken together suggest that more subtle factors such as root longevity, age-dependent nutrient absorption (Volder et al. 2005), or differences in root density (Southey and Archer 1988, Swanepoel and Southey 1989), in addition to the overall size of the root system, contribute to differences in scion performance when grafted onto a given rootstock (Pouget 1987).
Conclusions
We assembled numerous reports of trench wall profiles for grapevine roots from the literature and analyzed them according to an accepted method for characterizing root depth distribution. The analysis of the data using the parameter ß provides some comparative statistics on various rootstocks and on how soil characteristics may influence root distribution in the upper one to two meters of soil. The reports we reviewed show a general focus on genotype and cropping system differences as determinates of grapevine rooting depth distributions. As grapevine cultivation moves onto more marginal soils, there is a need for more information on how root distributions are influenced by diverse soil profiles, by infertile or shallow soils, and by unusual soil chemistry. There is a need for more conclusive information on whether or not characteristics such as emergence angles observed under greenhouse or laboratory conditions correlate to depth distribution in the field, and to understand which physiological, morphological, or demographic characteristics among diverse root-stock genotypes contribute to vine performance.
Footnotes
Acknowledgments: Support from The American Viticulture Foundation and the USDA/CSREES Special Research Grants Program 2001-34360-10328 and 2003-34360-13177 is gratefully acknowledged. The root mapping data in New York was obtained from unpublished maps done in 1962-63 provided for retracing and further analyses to A. Lasko by Dr. Nelson Shaulis of Cornell University, now deceased. We thank Anna Vu and Christine Stockert for technical assistance. David M. Eissenstat, Kerri L. Steenwerth, and M. Andrew Walker made many helpful comments that greatly improved both the scope and clarity of this manuscript.
This article was first published in the Proceedings of the Soil Environment and Vine Mineral Nutrition Symposium. L.P. Christensen and D.R. Smart (Eds.), pp. 153-169 (2005). American Society for Enology and Viticulture, Davis, CA.
From Proceedings of the Soil Environment and Vine Mineral Nutrition Symposium
- Copyright © 2006 by the American Society for Enology and Viticulture
Literature Cited
Vol 57 Issue 1








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
More from this TOC section
Similar Articles
