


Abstract
Grape phenolics are structurally diverse, from simple molecules to oligomers and polymers that are usually designated “tannins,” referring to their ability to interact with proteins. Anthocyanin pigments and tannins are particularly important for red wine quality. Their extraction depends on their location in the berry and their solubility. All phenolic compounds are unstable and undergo numerous enzymatic and chemical reactions. Color and taste changes during red wine aging have been ascribed to anthocyanin-tannin reactions. The structures and properties of tannins and pigmented tannins from these reactions are often misunderstood. Current research on wine phenolic composition is reviewed, with emphasis on the following issues: (1) reactions of tannins yield both larger polymers and smaller species; (2) anthocyanin reactions can generate colorless species as well as polymeric and small various pigments; (3) some polymeric pigments undergo sulfite bleaching while some low molecular weight pigments do not; (4) polymers are both soluble and astringent, so the astringency loss during aging may involve cleavage rather than polymerization; and (5) sensory properties of anthocyanins and tannins are modulated by interactions with other wine components.
Phenolic compounds are essential for the quality of plant-derived food products and wine. They are responsible for the color of red grapes and wines and are involved in the oxidative browning of white wines. They also contribute to taste and astringency through interactions with salivary proteins.
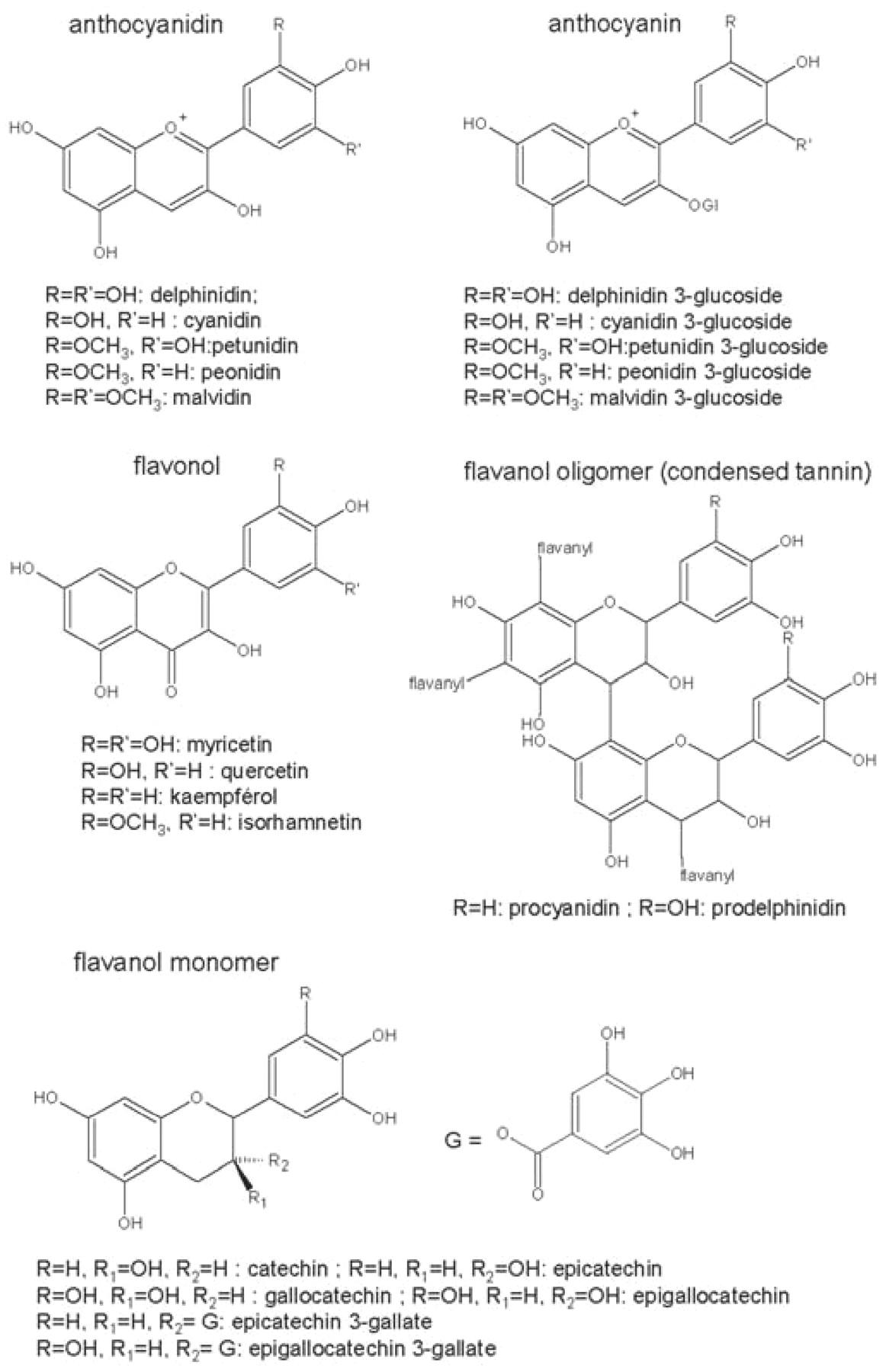
Two groups of phenolic compounds are classically distinguished: flavonoids based on a common C6-C3-C6 skeleton (Figure 1⇓) and nonflavonoids. Each group is further divided into several families, with shared structural features that confer specific properties such as color, aroma, and taste. Two groups of flavonoids, the anthocyanins and flavanols, are particularly important to the quality of red wines. Anthocyanins are the red grape pigments. Flavanols exist as catechin monomers and as oligomers and polymers, also called condensed tannins or proanthocyanidins. Other flavonoids are also present in low amounts, such as flavonols (for example, quercetin) and dihydroflavonols (for example, astilbin).

Structures of important flavonoids in grapes and wines.
Wine phenolic composition depends on the grape used and on winemaking processes that determine their extraction into the must and subsequent reactions. Anthocyanins and most tannins are localized in the solid parts of the cluster and are extracted by maceration in the fermenting must. They are highly unstable and undergo various enzymatic and chemical reactions as the wine is made and aged. Because the new compounds formed often exhibit sensory properties different from those of their precursors, these structural modifications change wine quality. In particular, the color change from the purple tint of young red wines to the tawny nuance of older ones is ascribed to reactions of grape anthocyanins with tannins generating new polymeric pigments (Somers 1971). Similarly, the decrease in astringency as wine ages results from polymerization of tannins and/or formation of polymeric pigments. The occurrence of such reactions in wine and their role in sensory changes is generally acknowledged, but the structures of the resulting products have only recently been established through new analytical techniques, and studies on their sensory properties have just started. Although color and taste depend primarily on molecular structure, they are modulated by interactions between each other and with other compounds present in the wine and in saliva. This review summarizes current knowledge on the structure of grape and wine pigments and tannins and on their role in determination of wine quality.
Structure of Anthocyanins and Tannins
Simple phenolic compounds and their distribution within the grape berry are well documented. Anthocyanins (Figure 1⇑) are specific to red varieties and localized in berry skins, except in teinturier varieties that have colored flesh. They are glucosylated derivatives of five aglycones or anthocyanidins: cyanidin, peonidin, petunidin, delphinidin, and malvidin. Further diversity results from acylation of the glucose by acetic, p-coumaric, and caffeic acids. Anthocyanin profiles are varietal characteristics and can be used to distinguish between them (Roggero et al. 1988). For instance, Pinot noir grapes have a very simple anthocyanin composition with only the five anthocyanidin 3-glucosides, while most other varieties also contain acylated derivatives. The proportions of acylated anthocyanins have been proposed for authentication of varietal wines, but they can be modified by winemaking processes such as use of pectolytic enzymes (Fischer et al. 2007) or maceration conditions (Canals et al. 2005), so this method is not reliable.
Flavanols exist in grape as monomers and as oligomers and polymers, called proanthocyanidins because they release red anthocyanidins when heated in acidic solutions. Proanthocyanidins show a great diversity of structures, from simple dimers to extremely complex molecules. They consist of several constitutive units that can be linked through different positions to make chains of variable length. Polymers with up to 80 units have been reported in grape skin tannins (Souquet et al. 1996). The complex structures of proanthocyanidins interact with and precipitate proteins involved in astringency perception. The designation “tannins” refers to this particular property.
Monomers and small oligomers can be analyzed by HPLC, but most grape flavanols are large oligomers and polymers (Czochanska et al. 1980, Souquet et al. 1996). Proanthocyanidin heterogeneity increases with chain length, resulting in poor resolution with all separation methods. Depolymerization methods such as thiolysis and phloroglucinolysis give access to the nature and proportions of constitutive units and enable calculation of mean degrees of polymerization (mDP). Grape seed proanthocyanidins are procyanidins (releasing cyanidin) based on catechin, epicatechin, and epicatechin 3-gallate units. Proanthocyanidins from other grape tissues also contain epigallocatechin units, and thus consist of both procyanidins and prodelphinidins (releasing delphinidin). Tannin extracts from skins have mean degrees of polymerization of ~30 (Souquet et al. 1996) compared with ~10 in extracts from seeds (Prieur et al. 1994) and stems (Souquet et al. 2000). The proportion of epicatechin gallate units is also quite different in skin (5%), stem (15%), and seed (30%) tannins. As a consequence, epigallocatechin units and epicatechin 3-gallate units are markers of proanthocyanidins from skins and seeds (plus stems), respectively.
Phenolic Composition from Grape to Wine
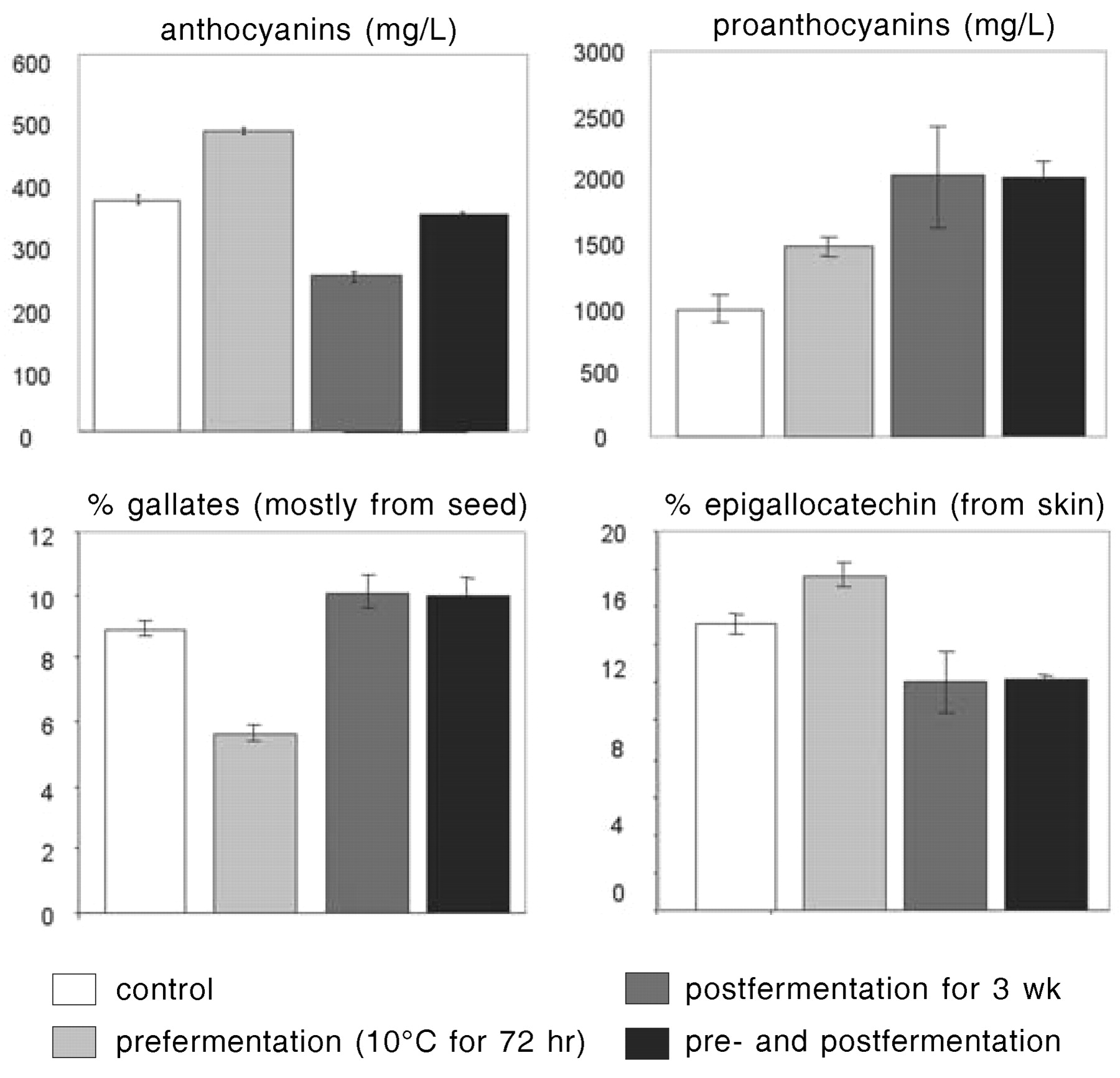
Phenolic changes associated with winemaking begin with selective extraction of grape constituents into the must during pressing or maceration. Hydroxycinnamic acids are the major phenolic compounds present in grape berry flesh and thus in white musts made by direct pressing of berries. Preparation of a red wine requires a maceration step to extract red pigments and tannins from the fruit. Diffusion of anthocyanins and skin proanthocyanidins into red grape must is faster than extraction of seed proanthocyanidins, which requires higher alcohol concentration and temperature (Cheynier et al. 1997). Monitoring of polyphenols extracted from skins or seeds in solutions with different ethanol concentrations showed that extraction of phenolics from seeds is accelerated by higher ethanol concentrations (Canals et al. 2005). Final levels of phenolics, especially p-coumaroylated anthocyanins and seed-derived procyanidins that are poorly water soluble, increased with ethanol concentration. In winemaking experiments, prefermentation maceration at low temperature increased concentrations of skin-derived anthocyanins and proanthocyanidins, whereas extended maceration beyond the end of alcoholic fermentation resulted in more proanthocyanidins from seeds and less anthocyanins (Figure 2⇓). Concentration of anthocyanins starts to decrease after a few days of skin contact, when the rate of their conversion to other molecular species exceeds that of their extraction (Cheynier et al. 1997). Some anthocyanins may readsorb to the grape solids or yeast, but anthocyanins recovered from pomace after pressing usually do not compensate for the losses measured from grape to wine.

Influence of pre- and postfermentation maceration on wine anthocyanin and proanthocyanidin composition.
Studies of phenolic reactions in wine involve two complementary approaches. New compounds can be isolated from wines and identified. Mechanisms for their synthesis are then postulated and checked in model wines containing potential precursors. Conversely, studies of model wines yield new products that can be searched for in wine.
Reactions of phenolics are discussed elsewhere (Fulcrand et al. 2006). However, the following points are worth emphasizing. Products formed from anthocyanins in the course of wine aging have been described as polymeric pigments resulting from reactions of anthocyanins with tannins. Several polymers resulting from direct reactions of anthocyanins with tannins (Salas et al. 2004a) or from reactions also involving aldehydes such as acetaldehyde (Timberlake et al. 1976) were actually detected in wines (Salas et al. 2005). However, some of these anthocyanin-tannin adducts are colorless (Remy et al. 2000). Moreover, anthocyanin reactions that do not involve tannins have also been demonstrated. The resulting pigments include both anthocyanin polymers (Salas et al. 2005, Vidal et al. 2004c) and small molecules such as pyranoanthocyanins (Cheynier et al. 1997, Fulcrand et al. 1996, 1998), also called vitisins (Bakker et al. 1997), that result from addition of yeast metabolites to anthocyanins and caftaric acid-anthocyanin adducts that are formed through enzymatic oxidation reactions (Sarni-Manchado et al. 1997). Finally, anthocyanin degradation reactions generate colorless, low molecular weight compounds (e.g., syringic acid) (Salas et al. 2003). Examples of anthocyanin derived structures known to occur in wine are shown in Figure 3⇓.

Anthocyanin derivatives detected in wine (R1, R2 = H, OH, OCH3).
Tannins yield higher molecular weight species through oxidation, polymerization, and addition with anthocyanins. However, in acidic-medium-like wine, they also undergo cleavage reactions (Haslam 1980, Vidal et al. 2002). Some of the cleavage products serve as intermediates in polymerization reactions, and some bonds in the new polymers are also labile. In particular, ethyl bridges formed through acetaldehyde-induced polymerization are unstable, and through cleavage and addition reactions are gradually converted to more stable pyranoanthocyanin derivatives (Cheynier et al. 1999). As a consequence, phenolic changes during wine aging are a dynamic process leading to increasing structural diversity but not necessarily to larger polymers. However, estimation of tannin mDP becomes increasingly inaccurate in older wines as some modified constitutive units (e.g., through oxidation) and other units incorporated into the polymers cannot be analyzed.
Structure/color relationships of phenolic pigments.
Absorbance spectrum in the visible range that determines color depends directly on the molecular structure and more specifically on the extent of electron delocalization within a given molecule. The flavonoid family exhibits a great diversity of colors, from colorless flavanols, through yellow flavonols and chalcones, to red and blue forms of anthocyanins, as further oxidation of the central heterocycle increases its conjugation with adjacent phenolic rings.
Anthocyanins are usually represented in the red flavylium cation form, the color of which shifts toward higher wavelengths (from orange to purple) as the number of substituents on the B ring increases. However, when dissolved in water, flavylium cations undergo proton transfer (Figure 4a⇓) and hydration (Figure 4b⇓) reactions, generating blue quinonoidal bases and colorless hemiketals (or carbinol), respectively. The proportion of each form is determined by pH, and flavylium ions predominate only in very acidic solutions (pH <2). At wine pH, grape anthocyanins occur mostly as the colorless, hydrated hemiketal form. In addition, sulfite ions react with the anthocyanin flavylium in the same way as water to form colorless products, through a reaction known as sulfite bleaching (Figure 4c⇓).

Reactions of anthocyanins in aqueous solutions and wine (R1, R2 = H, OH, OCH3): proton transfer (a); hydration (b); and sulfite bleaching (c).
Red wine color is ensured through two stabilization processes: conversion of grape anthocyanins to other pigments and association mechanisms collectively called copigmentation. Anthocyanin-derived pigments show a wide range of colors from orange to blue. Tannin-anthocyanin adducts are red, like their anthocyanin precursors (λmax 515 to 526 nm). Other reactions products include orange pyranoanthocyanins (λmax 480 to 510 nm) (Benabdeljalil et al. 2000, Sarni-Manchado et al. 1996), purple ethyl-linked species (λmax 528 to 540 nm) (Atanasova et al. 2002b, Timberlake et al. 1976), and even blue flavanyl-vinylpyranoanthocyanins (λmax 575 nm), called portisins since they were first isolated from port wines (Mateus et al. 2003).
These new structures may also change color with pH variations. Flavanol-anthocyanin adducts, like anthocyanins, are red only in very acidic solutions and are almost colorless at wine pH, indicating that hydration of the flavylium cation occurs (Salas et al. 2004b). No such bleaching is observed with flavanol-ethyl-anthocyanin pigments, which turn blue as pH increases. Spectrophotometry shows that the hydration constant of catechin-malvidin 3-glucoside is 2.64, like that of malvidin 3-glucoside, and no formation of quinonoidal base was observed at 1 < pH < 4. In contrast, the hydration constant calculated for catechin-ethyl-malvidin 3-glucoside was 4.17, and much higher than its proton transfer constant (3.44), meaning that it is mostly present in wine as the blue quinonoidal base. The constants for the first and second hydrations of malvidin 3-glucoside-ethyl-malvidin 3-glucoside, 1.81 and 4.63, respectively (Atanosova et al. 2002b), indicate that at wine pH, one of the anthocyanin moieties is in the red flavylium form and the other is hydrated.
Pyranoanthocyanins remain colored over a wide pH range and in the presence of sulfites (Bakker et al. 1997, Sarni-Manchado et al. 1996). That is attributed to the substituent on the C ring which impedes nucleophilic addition of water (Brouillard 1982) or sulfites (Timberlake et al. 1967).
Color stabilization may also take place through copigmentation, interactions that involve other molecules present in the medium (intermolecular copigmentation) or other parts of the pigment molecule itself (intramolecular copigmentation). The mechanism involved in copigmentation has been thoroughly studied (Brouillard and Dangles 1993, Goto et al. 1991). Colored anthocyanins (red flavylium or blue quinonoidal base) are planar structures that can interact with other planar species (copigments) to form molecular stacks from which water is excluded. The flavylium ion is thus trapped and protected from hydration, and the hydration equilibrium is displaced from the colorless hydrated form toward the red flavylium and then the blue quinonoidal base. This is particularly important at wine pH, where hydrated forms normally predominate. It increases color intensity (the hyperchromic effect) and shifts the color toward purple (the bathochromic effect). Studies using chlorogenic acid as the copigment have shown that the various anthocyanin derivatives are differently affected by copigmentation. For instance, at a pigment:copigment ratio of 1:100, there was a 42% increase in the visible absorbance spectrum of catechin-malvidin 3-glucoside. Under the same conditions, the absorbance of catechin-ethyl-malvidin 3-glucoside was not modified and that of malvidin 3-glucoside-ethyl-malvidin 3-glucoside underwent a slight bathochromic shift. That suggests the chromophore of ethyl-anthocyanin derivatives is already protected by intramolecular copigmentation or formation of sandwich-type stacks through self-association, as demonstrated for similar products obtained from a synthetic anthocyanin (Escribano-Bailon et al. 2001). These processes may also help ethyl-antho-cyanin derivatives resist sulfite bleaching.
Reactions of anthocyanins with tannins and other molecules yield a variety of structures with a wide range of molecular weights that include both pigmented and colorless species. Pigments resistant to sulfite bleaching are not necessarily polymeric, and some polymeric pigments are not resistant to sulfite bleaching. Thus, conversion of anthocyanins (75 to 95% colorless, 5 to 25% red at wine pH) to flavanol-anthocyanin polymers has no effect on color, but formation of pyranoanthocyanins (100% colored) and ethyl-linked derivatives (50% or more colored) shifts color from red to orange or purple tints and enhances color intensity and resistance to pH changes and sulfite bleaching. It should be emphasized that the chemical stability and color of pigments are not related. Ethyl-linked anthocyanins are less stable than anthocyanins in aqueous solution at wine pH because of acid-catalyzed cleavage of their ethyl bonds (Escribano-Bailon et al. 2001). Reactions of the cleavage products lead to flavanyl-pyranoanthocyanins, which change in color from red to tawny and increase resistance to sulfite bleaching (Atanasova et al. 2002a).
Relationships between phenolic structure and taste.
Phenolics may be bitter or astringent, and some are also volatile and contribute specific odors such as vanillin, ethyl phenols, and vinyl phenols. Two perception mechanisms must be distinguished: perception of taste (saltiness, sweetness, sourness, bitterness) and of odors involves interactions of the substance with taste or olfactory receptors, whereas perception of astringency is a tactile sensation elicited by precipitation of salivary proteins, which reduce mouth lubrication. Thus, bitterness is restricted to small molecules with particular structural features enabling them to enter the receptor and activate the signal transduction process, whereas astringency depends on the number of protein interaction sites in the molecule and increases with its size.
Within the flavanol series, bitterness decreases and astringency increases from monomer to trimer (Noble 1998), as expected from these mechanisms. Astringency of larger proanthocyanidin polymers (mDP 3 to 20) also increases with the molecular weight and the percentage of galloylation (Vidal et al. 2003), as does their ability to complex with peptides and proteins (Sarni-Manchado et al. 1999a,b, 2002). Astringency may be modulated by accessibility of interaction sites and thus molecular conformation, as demonstrated in protein-tannin interaction studies (Poncet-Legrand et al. 2006).
Changes in wine sensory parameters during aging are ascribed to tannin reactions, but studies on taste properties of derived tannins are scarce. Oligomers arising from catechin oxidation are likely to contribute astrigency like their procyanidin isomers and interact similarly with proteins (Guyot et al. 1996). Ethyl-linked catechin oligomers were as astringent and more bitter than procyanidins (Vidal et al. 2004b). Astringency of wine extracts correlated with the concentration of proanthocyanidin units, but the analytical method used could not distinguish between subunits present in tannins and those from other polymers (Brossaud et al. 2001). Grape anthocyanins and isolated polymeric pigment fragments from wine or pomace were not perceived as bitter or astringent when tasted in 5% ethanol (Vidal et al. 2004b). A similar hydrosoluble wine fraction was estimated to contain flavanol-anthocyanin copolymers (three flavanol units per anthocyanin on average) with traces of carboxypyrano-anthocyanins and anthocyanin oligomers (Salas et al. 2005). These results suggest that incorporation of anthocyanins into tannin structures may lead to a decrease in astringency.
Higher molecular weight proanthocyanidins (over 10 subunits) are commonly believed to be insoluble and thus unable to contribute astringency (Lea 1990). However, proanthocyanidin fractions isolated from apple (mDP 70) and grape (mDP 20) were both readily soluble in a winelike water-alcohol solution and highly astringent (Vidal et al. 2003). High molecular weight proanthocyanidins (mDP >20) were precipitated selectively from wines by proteins added as fining agents (Maury et al. 2001, Sarni-Manchado et al. 1999b), indicating they were present in wine, interact strongly with proteins, and consequently are presumed to be astringent. Thus, flavanol polymerization reactions, regardless of the polymers formed (proanthocyanidins, oxidation products, or ethyl-flavanols), enhance rather than reduce astringency. However, tannins also undergo cleavage reactions generating low molecular weight species that contribute to loss of astringency during wine aging. Finally, conversion of tannins to oligomeric tannin-anthocyanin copolymers reduces wine astringency, but the tastes of other polymers such as ethyl-linked tannin-anthocyanin polymers and tannin-pyranoanthocyanins remain to be investigated.
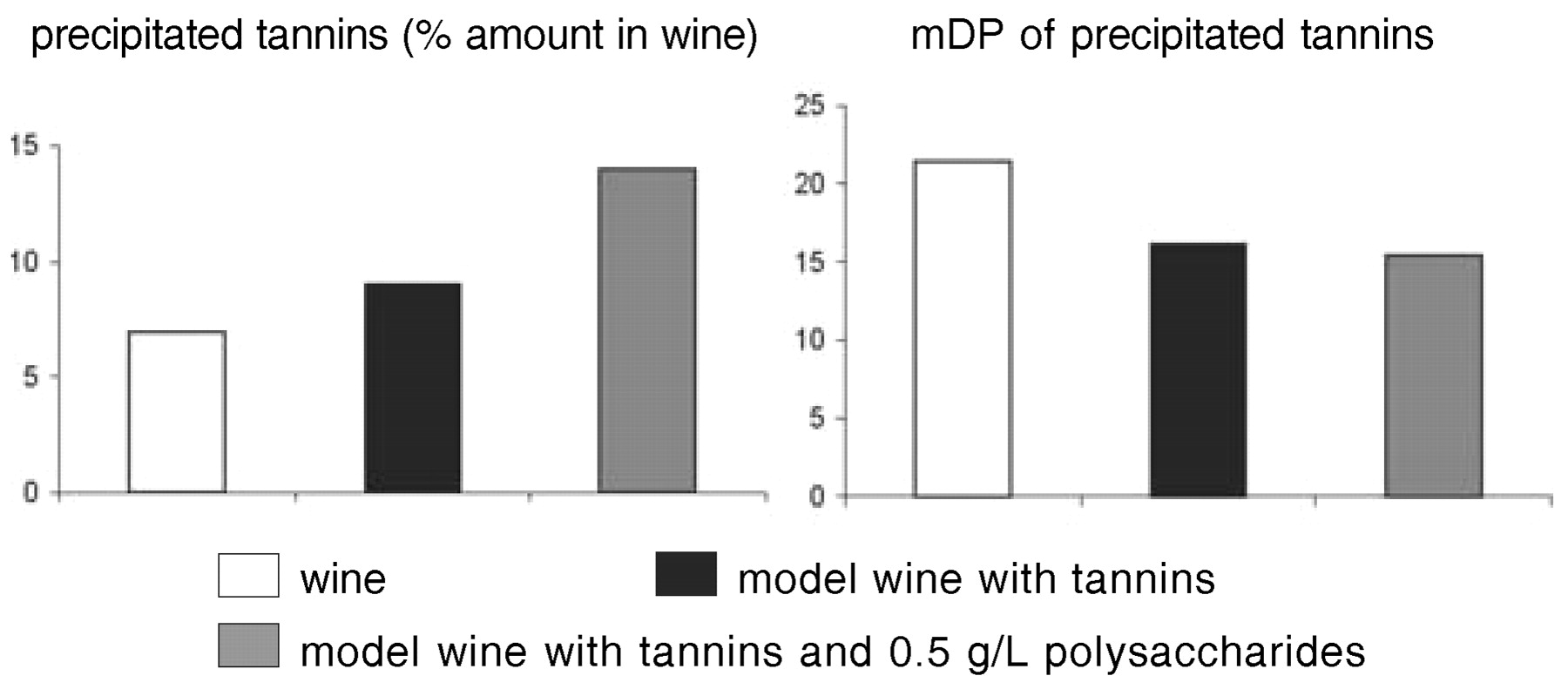
Beyond molecular structure, the tastes of phenolic compounds are modulated by interactions with other wine components such as ethanol, glycerol, salts, acids, and macromolecules. Fining with gelatin and other proteins selectively precipitates out high molecular weight tannins that are also particularly astringent, but the amount removed is generally too small to significantly alter wine composition (Maury et al. 2001). Some added protein remains in the wine to form soluble or colloidal complexes with wine tannins. Incorporation of the tannins in such complexes may impede their interaction with salivary proteins and thus reduce astringency. Tannins also interact with some wine polysaccharides to yield soluble complexes (Riou et al. 2002), which may have similar effects. Precipitation of wine tannins by fining proteins was higher in model wine prepared by dissolving tannins in ethanol-tartaric acid solution than in the original wine (Figure 5⇓). Adding polysaccharides at wine concentrations restored the initial precipitation rate, indicating competition between polysaccharides and proteins for interaction with tannins or formation of soluble protein-polysaccharide-tannin complexes (Maury 2001). Polysaccharides also reduced the selectivity of gelatin for higher molecular weight tannins. Thus, the quantity and quality of tannins precipitated by a given amount of protein depends not only on the ratio of tannins to proteins (Sarni-Manchado et al. 1999) but also on the wine matrix composition.
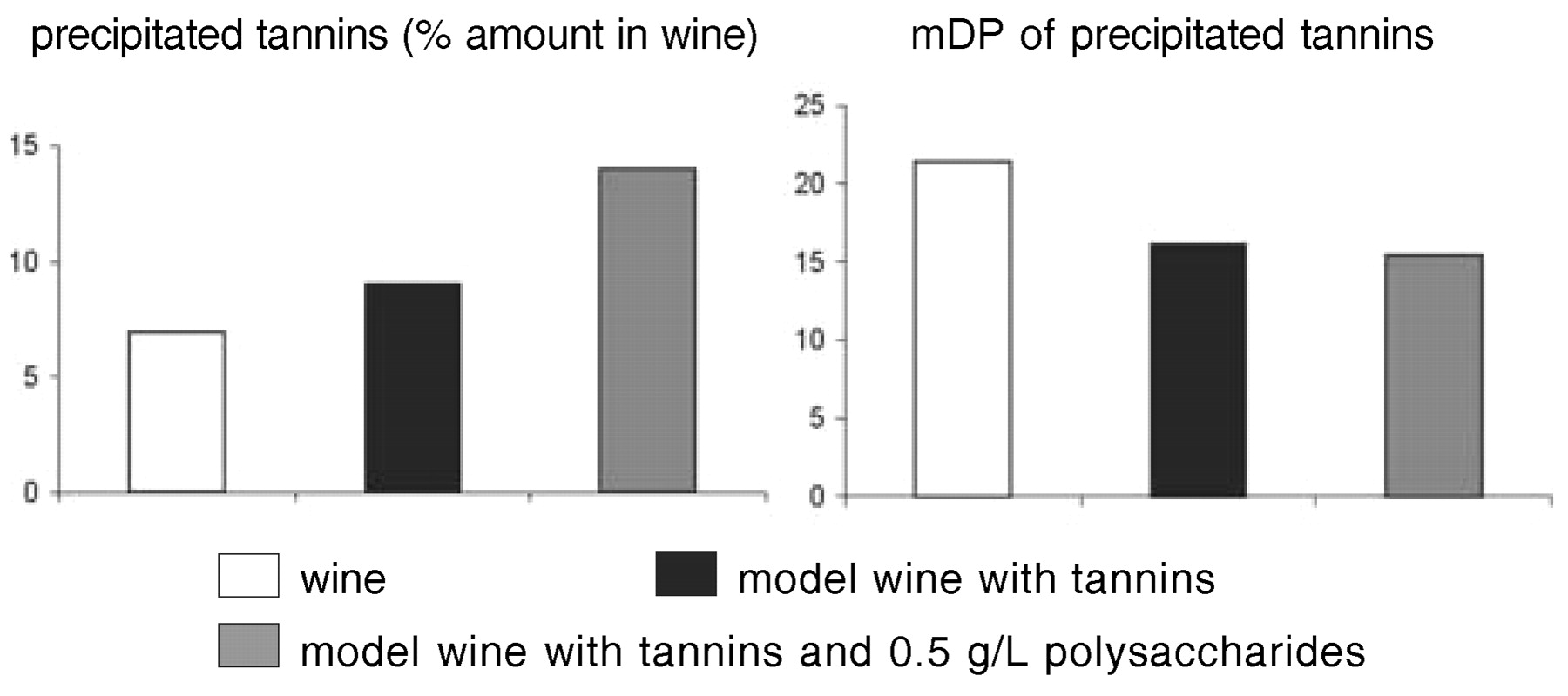
Influence of polysaccharides on tannin precipitation by gelatin fining.
Tannins also form colloidal particles in wine. Their size increases with tannin concentration, degree of polymerization, extent of galloylation, wine acidity, and wine ionic strength. Their size decreases as ethanol increases (Poncet-Legrand et al. 2003). All factors that promote aggregation also increase the intensity of astringency sensation as measured by sensory analysis (Vidal et al. 2004a). However, the relationship between aggregation and astringency is not straightforward, as addition of mannoproteins decreased tannin aggregation but had no effect on astringency, whereas addition of another polysaccharide, RGII, increased aggregation but decreased astringency.
Conclusions
New analytical techniques have unraveled some structures derived from tannins and anthocyanins in wine and determined how they are formed. Each species is present in very small amounts; even combined they are a small proportion of total wine phenolics with a limited contribution to wine color. However, most may be regarded as markers for many related structures similarly formed from the complete series of grape anthocyanins and tannins. Furthermore, some derivatives are unstable and undergo cleavage and polymerization reactions, leading to an increased diversity of structures. Despite recent findings, determination of wine phenolic composition remains a major challenge of wine research. However, availability of marker molecules representing some of groups of derived products has made it possible to study their color and taste properties.
Color stability with respect to pH changes and sulfite bleaching is not related to molecular weight, as polymers resulting from tannin anthocyanin reactions behave exactly like their anthocyanin precursors. The color resistance of orange pyranoanthocyanin pigments is due to their 4-substituted structure, whereas that of purple, ethyl-linked anthocyanin derivatives results from molecular stacking.
Astringency is related to tannin concentration and increases with their molecular weight, without size limit. Thus, loss of astringency during wine aging results from cleavage reactions and incorporation of anthocyanins into small tannin structures rather than from polymerization reactions. Other wine components such as polysaccharides and proteins interfere with astringency perception, presumably through competition with salivary proteins in the formation of tannin complexes.
The physicochemical basis for taste perception associated with tannins remains to be established. Hard tannin perception is associated with high tannin concentrations, especially of large polymers and galloylated tannins from seeds that aggregate easily. It is uncertain whether soft tannin perception is due to low concentrations of such tannins, to the presence of specific compounds such as anthocyanin-tannin adducts, or to the incorporation of tannins into protein or polysaccharide complexes. Finally, the role of psychological factors should not be overlooked. In particular, green tannin perception appeared linked with lack of red color associated with poor ripeness. Green versus soft tannin perception was changed when the same series of wines was tested in white or black glasses by an expert wine panel (Brossaud 1999), indicating mental associations between color and taste perception.
Footnotes
Acknowledgments: Part of this research has been funded by Société Martin Vialatte, by grants from Consejo Nacional de Ciencia y Technologia (Mexico), Société Francçaise d’Exportation des Ressources Educatives (France), and Ministerio de Educacion, Cultura y Deporte (Spain).
This article was originally presented at the ASEV 56th Annual Meeting Phenolics Symposium, 20–21 June 2005, Seattle, WA. All phenolics symposium articles were peer reviewed by two fellow presenters, and James Harbertson, Mark Downey, and Sara Spayd served as technical editors of the articles.
From the ASEV 2005 Phenolics Symposium
- Copyright © 2006 by the American Society for Enology and Viticulture
Literature Cited
Vol 57 Issue 3








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
More from this TOC section
Similar Articles
