


Abstract
The plant hormone abscisic acid (ABA) is a signaling molecule involved in grape berry development and ripening. To obtain a more comprehensive understanding of ABA synthesis and catabolism in developing grape berries, ABA and catabolite profiles of Merlot grapes grown in a commercial vineyard in the southern Okanagan Valley in British Columbia, Canada, were measured using HPLC–MS/MS in multiple-reaction monitoring mode. While pulp and skin of preveraison berries contained low to moderate concentrations of ABA, these tissues had high concentrations of the ABA catabolite dihydrophaseic acid (DPA), indicating that large quantities of ABA had been produced and catabolized early in berry development. In the pulp and skin, two peaks of ABA were found, the first at two weeks preveraison and the second at late veraison, which was followed by a decline in ABA as the berries ripened. Abscisic acid glucose ester (ABA-GE) in the pulp and skin rose just before veraison, and then declined after veraison. In developing seeds similar trends were observed, but with initial high levels of both ABA and DPA and with ABA-GE remaining high into ripeness. ABA applied to berries at veraison had a half life of one week and resulted in a 69% increase in DPA concentration. However, this constituted only a small increase in the total ABA and catabolite pool. The treatment had no effect on berry maturation rate, basic juice composition, or skin and seed tannins, but did increase the anthocyanin content of skins by 7% at berry maturity. These observations indicate that little ABA was taken into the berry and metabolized, while the majority of it remained on the surface.
Growth and ripening of fruit, including grape berries, is controlled by plant hormone classes such as ethylene, auxins, brassinosteroids, and abscisic acid. Abscisic acid ((S)-cis-abscisic acid, the specific isomer of this molecule that is naturally synthesized in plants, referred to hereafter as ABA; Figure 1A⇓), in particular, is involved in the production of anthocyanins in the skin of ripening berries (Ban et al. 2003) and is also produced in the roots and leaves of drought-stressed vines, regulating stomatal aperture and transpiration (Stoll et al. 2000). Studies in numerous other species, including Brassica, Arabidopsis, conifers, and lettuce, have shown that ABA is rapidly turned over in plants (Feurtado et al. 2004, Huang et al. 2008). For this reason, measurements of ABA concentrations alone give limited information on the total amount of ABA produced over time during development and on the potentially active products resulting from ABA catabolism within plant tissues. This study, which profiles ABA and catabolites over the course of berry development, was undertaken to provide baseline data on ABA biosynthesis and catabolism in Merlot grapes, with the ultimate aim of providing researchers with tools for developing strategies to optimize berry quality.

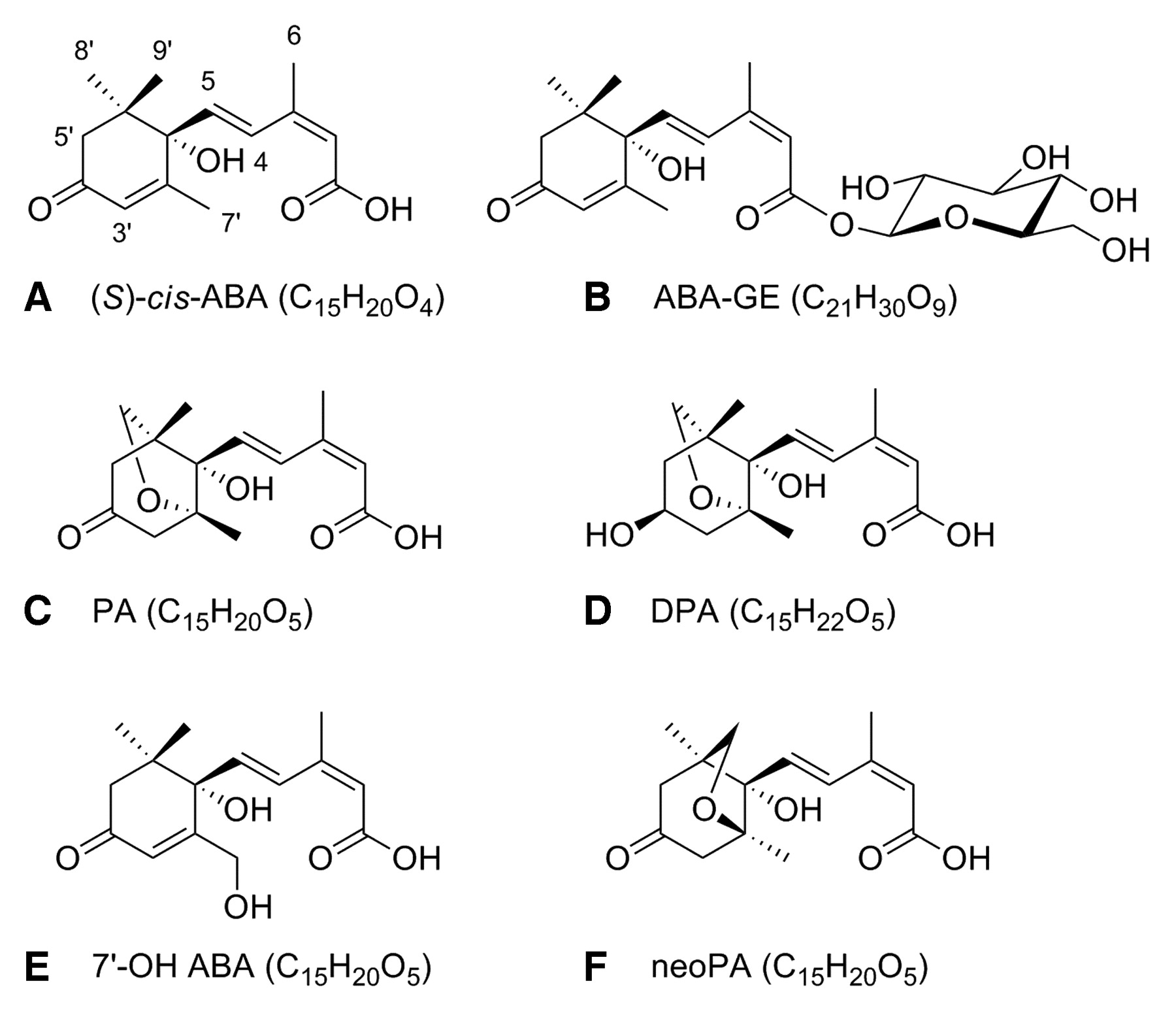
Structures of ABA and its five catabolites.
Abscisic acid has been shown to be involved in the regulation of primary and secondary metabolite biosynthesis during grape berry ripening (Antolín et al. 2003, Cawthon and Morris 1982, Davies et al. 1997). In a recent study of developing Merlot grape berries, ABA peaked at veraison and then declined (Deytieux-Belleau et al. 2007). Increases in ABA levels during berry maturation have been correlated with increases in soluble solids, reducing sugars, and anthocyanins. Effects have also been shown on the expression of genes involved in anthocyanin biosynthesis, invertase activity, and ASR (ABA, stress, and ripening) proteins involved in sugar metabolism and ripening (Ban et al. 2003, Çakir et al. 2003, Gagné et al. 2006, Pan et al. 2005). ABA has also been associated with environmental effects on berry ripening and anthocyanin accumulation, including temperature (Mori et al. 2005, Koshita et al. 2007), light (Jeong et al. 2004), and water stress (Antolín et al. 2006, Stoll et al. 2000).
Several studies have found positive effects of ABA applications on grape berry development and production of components associated with maturation and ripening. ABA applied to Flame Seedless grapes led to increases in berry size and reductions in juice acidity (Peppi and Fidelibus 2008) and to decreases in berry firmness and increases in skin anthocyanins (Peppi et al. 2006, Peppi and Fidelibus 2008). Anthocyanin content was also increased by applying ABA to Cabernet Sauvignon berries (Gagné et al. 2006, Pirie and Mullins 1976). In Kyoho, ABA enhanced anthocyanins and the expression of seven genes in the anthocyanin biosynthetic pathway (Ban et al. 2003) and activated cell-wall-bound and soluble acid invertases (Pan et al. 2005). Earlier research found that, in Thompson Seedless berries, ABA stimulated the gluconeogenic process and promoted protein synthesis (Palejwala et al. 1985).
The concentration of active forms of ABA in plant tissue depends upon the relative rates of biosynthesis, catabolism, and transport. ABA metabolism has been studied in-depth in numerous plant species (Zaharia et al. 2005). The biological role of conjugation and catabolism of ABA and the effects of ABA catabolites in ripening grape berries have not been studied to date. The principal pathway of ABA catabolism is by oxidation of the 8′-carbon atom, affording 8′-hydroxy ABA (8′-OH ABA), which can reversibly cyclize to phaseic acid (PA; Figure 1C⇑) (Zaharia et al. 2005). In some plant cells and tissues PA accumulates, while in other tissues dihydrophaseic acid (DPA; Figure 1D⇑), a product of further reduction, predominates. Competing oxidation pathways at different ring methyl groups of the ABA molecule also create the products 7′-hydroxy ABA (7′-OH ABA, Figure 1E⇑) and 9′-hydroxy ABA (9′-OH ABA). The latter can cyclize reversibly to neophaseic acid (neoPA; Figure 1F⇑). In the few biological systems in which the catabolites have been tested, the hydroxylated metabolites are biologically active while the ring-closed forms are not (Zou et al. 1995). ABA is rapidly turned over by plant enzymes, either by oxidation or by conjugation to abscisic acid glucose ester (ABA-GE; Figure 1B⇑). It has been proposed, but not demonstrated, that ABA-GE acts as a reservoir of ABA in grape berries (Kondo and Kawai 1998). Recently it was shown that a β-glucosidase in Arabidopsis was able to hydrolyze ABA-GE to ABA, suggesting that ABA-GE could represent an inactive storage form of ABA in plants (Lee et al. 2006). The glucose ester of ABA has not been analyzed directly during grape development, although ABAGE concentrations in Cabernet Sauvignon and Pione grape berries have been estimated by measuring levels of ABA after hydrolysis and subtracting the contribution of free ABA (Kondo and Kawai 1998, Koussa et al. 2004). In Cabernet Sauvignon berries, the concentration of free ABA in the skin and pulp was highest at half veraison, while the greatest accumulation of the conjugate was found at the end of veraison (Koussa et al. 2004). Later in maturation, the levels of both decreased, with the conjugate remaining at higher levels than the free acid. There are no reports in the literature of the occurrence of oxidation products of ABA in grape berries. The decline observed in both ABA and conjugated ABA in Pione grape berries later in maturation could be due to catabolism through oxidation (Kondo and Kawai 1998).
We modified an existing HPLC–MS/MS method, used for analyzing ABA and metabolite regulation of seed development and dormancy breaking in Brassica, Arabidopsis, and conifer seeds (Feurtado et al. 2004, Ross et al. 2004, Zhou et al. 2003), for use with grape berries (Owen and Abrams 2009). We report herein the natural profile of ABA and ABA catabolites in Merlot grape berries over the growing season and show the effects of treating berries at veraison with ABA on the profiles of ABA and catabolites and on fruit composition.
Materials and Methods
Field experiments.
The studies were conducted during 2004 and 2006 on 6-year-old Merlot vines (clone 347 on 3309 rootstock) planted on an Osoyoos loamy sand at Vincor Canada’s Sunrock Vineyard, located 6 km southeast of Oliver, British Columbia, Canada. The trellised rows of vines were m apart with m in-row vine spacing. The vines were bilateral-cordon trained and thinned before bloom each year to retain 16 shoots, each bearing one cluster. Drip irrigation was applied 1 to 2 times weekly when the volumetric soil moisture content to 60 cm depth, measured using a capacitance probe (C-Probe, Adcon Telemetry, Boca Raton, FL) installed in the vineyard block, was depleted to below 10%.
In 2004, three replicate samples, containing five berries each, of immature and mature berries were collected from separate vines in different vineyard rows. The berries were sampled from the midregion of separate clusters on five randomly selected shoots per vine. The immature berries were sampled on 11 Aug at midveraison. The mature berries were sampled on 23 Sep at commercial maturity. The samples were immediately frozen in liquid nitrogen and then stored at −80°C before shipping overnight on ice to the Plant Hormone Profiling Lab at the Plant Biotechnology Institute of the National Research Council (PBI-NRC) in Saskatoon, Saskatchewan, Canada. Here, they were divided into portions containing skins, pulp, and seeds for subsequent ABA and catabolite analysis.
In 2006, an experiment was conducted to study the changes in ABA and catabolite levels in berries over development and the effects of applying ABA to clusters at veraison. Two treatments, a control and an application of ABA, were applied to all clusters on single vines in a randomized complete block design with four replicates in separate vine rows. The ABA solution was prepared by dissolving 250 mg ABA in 1 mL 95% ethanol and bringing the solution to 1 L with water. A surfactant (Agral 90; Norac Concepts Inc., Guelph, Ontario, Canada) was added at 1 mL/L. The ABA solution was spray-applied on 1 Aug at 10 mL per cluster, ensuring that all berry surfaces were covered. A water/ethanol/surfactant spray was applied similarly to the clusters on control vines. Berries were sampled for ABA analysis biweekly from one month preveraison on 4 Jul to berry maturity on 26 Sep, and more frequently just before and during veraison including on the day of but prior to applying the ABA treatment, and at 1, 3, and 7 days after the ABA treatment. From each vine 10 berries were collected from the midregion of separate clusters and immediately frozen in liquid nitrogen for ABA analysis. The frozen samples were stored at −80°C before shipping to the Plant Hormone Profiling Lab, as described above, for analysis.
Sampling and analysis for fruit composition.
Berries were sampled from each vine for basic compositional analysis on 12 and 26 Aug and 23 Sep in 2004, and on 30 Aug and 26 Sep in 2006. The 20-berry samples consisted of two berries from the midregion of the exposed south-facing side of each of 10 clusters per vine. The fresh berries were crushed with a mortar and pestle before the juice was pressed through cheesecloth. Juice soluble solids (SS) content was measured using a digital refractometer (PR-101; Atago, Japan). Titratable acidity (TA) was measured by titration with N NaOH to a pH endpoint of using an autotitrator (655 Dosimat, Metrohm, Switzerland). Juice pH was measured using a standard pH meter (PHM82; Bach Simpson, Ontario, Canada).
Each year at harvest maturity, a 100-berry sample was collected randomly from among all clusters on each vine and stored at −30°C before the analysis of phenolics. The skins and seeds were removed, weighed, lyophilized, and ground in a mortar with liquid nitrogen prior to phenolics extraction. Phenolics were extracted from 250 mg of ground skin or seed three times, each using 8 mL of MeOH:H2O:HCl in the following ratios: 50:49:1, 75:24:1, and 99:0:1 v/v. The extracts were combined, centrifuged, and made up to 25 mL volume. Anthocyanins were determined using the Glories method (Glories 1984). Tannins were determined using a vanillin assay (Burns 1971).
ABA and catabolite analyses.
Berry samples collected for ABA and catabolite analysis were dissected, while frozen, into tissue fractions for separate analysis. Samples collected in 2004 were dissected into seed, skin, and pulp fractions, and those collected in 2006 were dissected into a combined skin and pulp (skin/pulp) and a seed fraction. The sample fractions were processed and analyzed for hormones according to a published method (Owen and Abrams 2009). Briefly, the method used was as follows. All sample fractions were first lyophilized and then homogenized with 10 -mm stainless-steel beads for 2 min using a Mini-Beadbeater-96 (BioSpec Products, Bartlesville, OK). A mixture of deuterium-labeled internal standards (IS) was added to 50 mg of each dried fraction, together with 3 mL of extraction solvent (IPA:H2O:AcOH, 80:19:1). The solution was vortexed and allowed to extract for 18–24 hr while shaken at ~380 rpm on an orbital shaker placed in a ~4°C refrigerator. After extraction, the samples were vortexed again, centrifuged, and then the supernatant was transferred to a new vial. This extract was dried on a Büchi Syncore Polyvap (Büchi, Switzerland) at 30°C and 340 rpm, and under vacuum reduced from 130 to 30 mbar over 1 hr, after which it was held at 30 mbar until dry. The dried extract was subjected to cleanup by mixed-mode cation exchange solid-phase extraction (Oasis MCX SPE cartridges, 3 cc, 60 mg; Waters Corporation Canada, Mississauga, ON). This step was included to eliminate the majority of the anthocyanins present in the berry extracts. The dried extracts from this step were then readied for a second stage of SPE using hydrophilic-lipophilic balance cartridges (Oasis HLB, 1 cc, 30 mg; Waters Corporation). This final extract was dried down in a centrifugal concentrator and redissolved in 200 μL of the reconstitution solvent before HPLC–MS/MS analysis.
All samples were analyzed using a Waters Alliance model 2695 HPLC coupled to a Micromass Quattro Ultima quadrupole tandem (QqQ) mass spectrometer (Waters Corporation) via a Z-spray electrospray (ES) ion-source. Both the HPLC and the mass spectrometer were controlled by MassLynx v software (Waters Corporation). HPLC conditions included a Genesis C18 analytical column ( × 100 mm, 4 μm; Chromatographic Specialties, Brockville, ON, Canada) with an Opti-Guard C18 guard column (1 mm, Optimize Technologies, Oregon City, OR), a column temperature of 34°C, an injection volume of 10 μL, and solvents A, acetonitrile, B, milli-Q water, and C, 5% aqueous acetic acid to create a gradient as shown (Table 1⇓). Mass spectrometry was carried out using negative electrospray ionization (ESI-) and multiple reaction monitoring (MRM), with a capillary potential of 0 to 5 kV, a source temperature of 120°C, a desolvation temperature of 350°C, cone gas and desolvation gas flow rates of 100 and 650 L/hr, respectively, quadrupole low and high mass resolutions of 1, a collision (Ar) gas pressure of e-3 mbar, and MRM transitions and conditions as shown (Table 2⇓). Analyte quantification was performed using QuanLynx v software (Waters Corporation) and internal standard-based calibration curves were prepared for each analyte/IS pair, as described elsewhere (Ross et al. 2004).
HPLC gradient conditions.
Multiple reaction monitoring (MRM) transitions and conditions used for mass spectrometry. The interchannel delay time was set to 4 seconds, while the interscan delay time was set to second.
Statistical analysis.
Effects of the ABA treatment on berry mass and composition and on ABA and its catabolites in skin/pulp at each posttreatment sampling date were determined by analysis of variance (ANOVA) conducted using the SAS GLM procedure (SAS Institute, Cary, NC).
Results
Fruit phenology and development.
Fruit phenology and time of maturation were typical for Merlot grown commercially in the region. In 2004, bloom (50% of flowers open) occurred on 8 Jun, veraison was from 7 to 17 Aug, and the fruit was considered mature on 23 Sep, when the berry flavor and aroma and the juice soluble solids and acids were at desirable levels for commercial harvest. In 2006, bloom occurred on 13 Jun, veraison was from 3 to 15 Aug, and the fruit was considered mature on 26 Sep.
ABA and catabolite profiles in berry fractions.
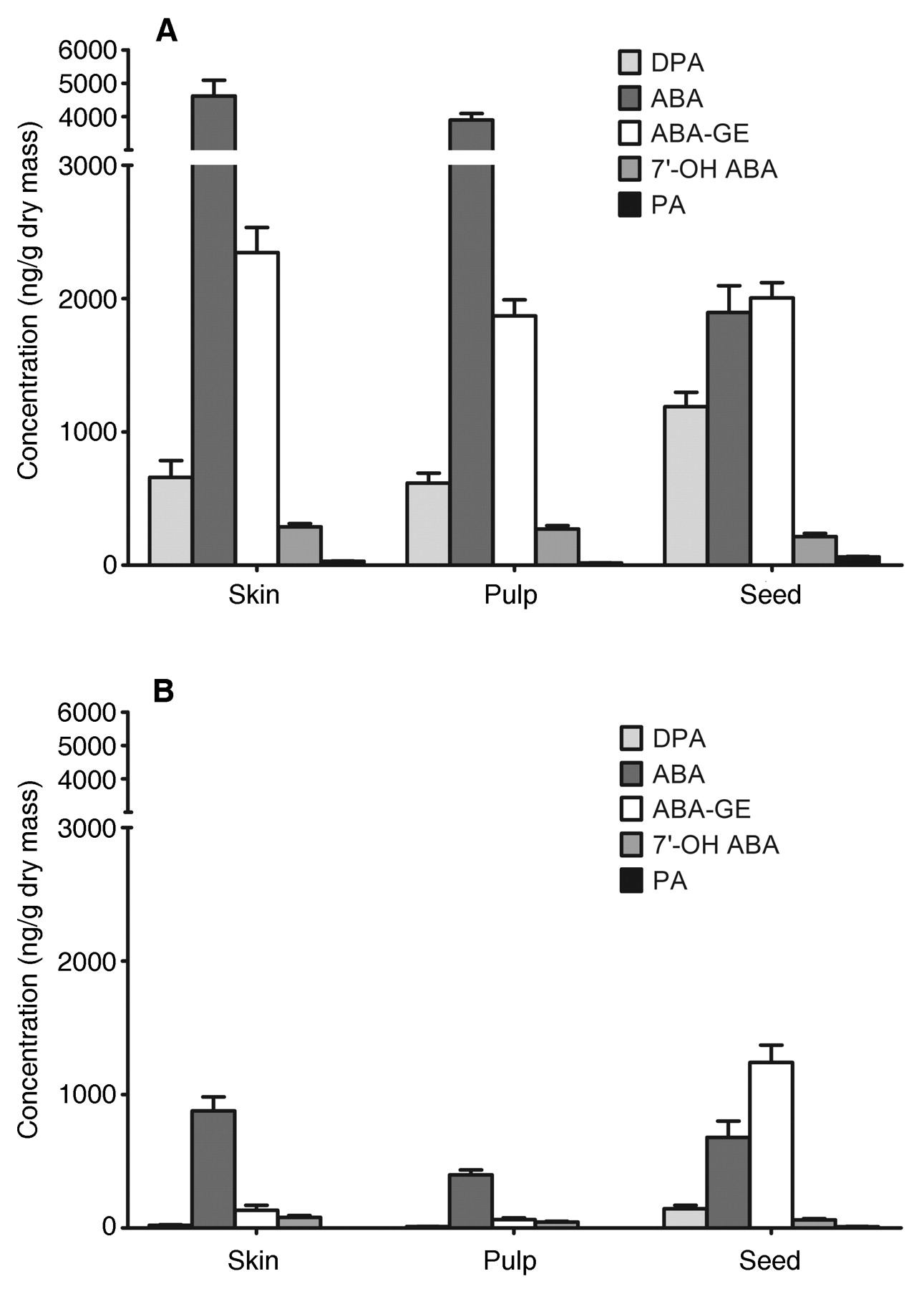
The profiles of ABA compounds in separated skin, pulp, and seeds in 2004 revealed that concentrations of the hormone and its catabolites in the skin and pulp were drastically reduced between midveraison and maturity (Figure 2A, B⇓). There was little difference between skin and pulp in the level of any compound at either stage of development, but there were substantial differences in the proportions of compounds between skin or pulp and seeds. At midveraison the ABA was much lower and the DPA higher in the seeds than in the skin or pulp. At maturity the ABA-GE was much higher in the seeds than in the skin or pulp. As well, the reduction of ABA and ABA-GE from midveraison to maturity was less drastic in seeds than in skin or pulp, such that at harvest maturity ABA levels were similar among tissues.

ABA and catabolite profiles of skin, pulp, and seeds from Merlot grapes. (A) Immature berries harvested at midveraison, ~4 days after the onset of veraison, on 11 Aug 2004. (B) Mature berries harvested at commercial maturity ~7 weeks after the onset of veraison, on 23 Sep 2004. Error bars indicate the standard error of the mean of three biological replicates.
ABA and catabolite profiles over the growing season.
At midveraison and harvest maturity, which were the sampling times common between 2004 and 2006, levels of ABA and catabolites were similar between the years except that higher levels of ABA-GE were found in skin/pulp and seed at maturity in 2006 (Figure 2A, B⇑; Figure 3A, B⇓). Over berry development from preveraison to full maturation in 2006, there were two ABA peaks in skin/pulp: on 19 Jul, two weeks preveraison, and on 15 Aug, at late veraison (~80% colored berries; Figure 3A⇓). The most striking feature of the profiles for both skin/pulp and seed was the high level of dihydrophaseic acid (DPA) at the earliest sampling dates, 4 and 19 Jul. In early berry development, ABA turnover appears to proceed through the oxidation pathway.

ABA and catabolite profiles of Merlot grapes over the growing season in 2006: (A) skin/pulp, (B) seeds, and (C) skin/pulp of berries treated with ABA on 1 Aug. Asterisks (*) in C indicate the treatment mean is significantly different from the corresponding control mean in A. Error bars indicate the standard error of the mean of four biological replicates.
The ABA-GE concentration increased two-fold from 4 Jul to 1 Aug. In skin/pulp it remained stable through midveraison and then declined slowly. In ripe berries the skin/pulp contained relatively low levels of ABA and the glucose ester. Levels of neoPA were below the limit of quantification (LOQ) in all samples at all dates.
The grape seeds exhibited mostly the same trends as the skin/pulp over the growing season but with notable exceptions (Figure 3B⇑). At the earliest sampling dates 2 to 4 weeks before veraison, ABA was much higher in seeds than in skin/pulp. The difference was ~six-fold at the earliest sampling on 4 Jul. Later, from late veraison to maturity, ABA levels declined less in seed than in skin/pulp, and ABA-GE remained elevated in seeds even at full berry maturity rather than declining as it did in the skin/pulp.
ABA supplied to grape berries.
In berries treated with ABA at the onset of veraison in 2006, ABA levels in and on skin/pulp rose dramatically by 2 Aug, the day after treatment, and by 15 Aug had declined to normal levels (Figure 3C⇑). A 69% increase in DPA was detected in the skin/pulp at 3 days posttreatment, but constituted only a small increase in the ABA catabolite pool. This increase was after normal DPA levels had declined drastically from the very high preveraison levels. By 7 days posttreatment, DPA had returned to typical low levels. There was no significant concomitant change in the profiles of the other ABA catabolites. Natural ABA is isomerized by light to trans-ABA, a nonnatural product, and the levels of this compound followed the same trend as natural ABA, but delayed by several days (data not shown).
Fruit composition parameters.
The size and basic juice composition of the mature berries were similar between 2004 and 2006 and typical for Merlot produced in the region. The berries averaged ~ g, with soluble solids of ~25 Brix, pH of ~5, and titratable acidity of ~ g/L (Table 3⇓). Juice composition from premature (30 Aug) and mature (26 Sep) berries was unaffected by the ABA application in 2006, which indicates that ABA did not affect the timing or rate of ripening as indicated by these parameters.
Mass and basic composition of Merlot berries harvested at midveraison on 12 Aug 2004 and at one month before and at maturity on, respectively, 26 Aug and 23 Sep 2004, and 30 Aug and 26 Sep 2006. Mean ± SE based on eight samples of 20 berries.
Skin and seeds, the phenolic-containing components of berries, together comprised 1% and 1% of the fresh mass of mature berries in 2004 and 2006, respectively (Table 3⇑, Table 4⇓). Skin, which contains the berry anthocyanins, comprised a higher proportion of the berry fresh mass in 2006 than in 2004 (11% versus 8%). Skin and seed fresh and dry mass and tannin contents were unaffected by the ABA treatment applied in 2006, but skin anthocyanin content increased by 7% on a skin dry mass basis and 6% on a per berry basis (Table 4⇓).
Characteristics of phenolic-containing components of Merlot berries in 2004 and 2006. Mean ± SE based on four samples of 100 berries.
Discussion
Plant hormones play important roles in the ripening of grapes and other fruit. In a comprehensive review on the molecular chemistry and biochemistry of development and ripening in fruit, researchers describe how different hormones regulate specific processes at different stages through complex signaling pathways not yet understood (Srivastava and Handa 2005). Increases in brassinosteroid levels have been correlated with ripening of grapes (Symons et al. 2006). Auxin has been shown to retard ripening and acts antagonistically to ABA, while ABA has been shown to promote maturation after veraison (Davies et al. 1997, Ban et al. 2003, Deytieux-Belleau et al. 2007).
Because ABA is turned over rapidly, quantifying ABA alone, or with its catabolites at a single stage of development, does not fully explain ABA synthesis and catabolism in the plant tissue. This fact is illustrated by the data collected for the skin/pulp (Figure 3A⇑) and seeds (Figure 3B⇑) in untreated berries. It is not evident from examination of the ABA concentration alone that ABA had been synthesized and turned over early in grape development. Only by quantifying the concentration of DPA in the sample does this usage become clear. Our method of quantifying ABA and its catabolites over the course of development allows for a wider survey of hormone catabolism.
In the Merlot berries analyzed in this study, the quantity and distribution of ABA changed over development. Concentrations of ABA and all catabolites were higher at veraison than when the berries were commercially mature. Skin and pulp had similar levels of all compounds at both veraison and maturity, whereas the proportion of ABA was higher in skin and pulp than in seed of immature berries, and ABA-GE was higher in seed than in skin and pulp of mature berries, suggesting that the ABA catabolism is different in the seed versus soft tissues. Similar findings were reported for ABA and ABA-GE (measured by difference, by hydrolysis of conjugates) in seeds of developing Cabernet Sauvignon grapes (Koussa et al. 1993). This pattern is not common to all grape varieties, however. In Pione grapes, seed ABA increased over berry development and conjugated ABA also increased but at a much lower level (Kondo and Kawai 1998).
In 2006, we observed that early in berry development (4 Jul) levels of the ABA oxidation product, DPA, in the skin/pulp were 10-fold higher than that of ABA and five-fold higher than the greatest concentration of ABA, which occurred at late veraison (Figure 3A⇑). A similar observation was made in a study of ABA catabolism in sweet cherry (Setha et al. 2005), and the authors suggested that the increased ABA catabolite concentrations at the early stage of cherry fruit development resulted from a lack of need for ABA. However, it can equally be argued that higher levels of ABA are needed for development and responses to environmental stress at the early stages and that the ABA is catabolized after action. This area needs further investigation.
The ABA concentration in the developing berries increased beginning at the onset of veraison, as has been found in other studies. In the lag phase between early development and veraison, it appears from the concentrations of the ABA catabolites that newly formed ABA was catabolized by conjugation to form ABA-GE and not by oxidation leading to DPA. The ABA-GE may act as a storage form for later release of ABA under stress conditions, as has been proposed for stress-induced release of ABA from ABA-GE in Arabidopsis (Lee et al. 2006). Alternatively, the hydroxylated catabolites of ABA may be active in some hormonal processes, as has been found for triacylglyceride synthesis in developing oilseeds (Zou et al. 1995). In this study ABA-GE levels in skin/pulp and seeds at maturity were substantially higher in 2006 than in 2004, whereas ABA levels were similar between years. The berries had similar total mass and seed mass between years, but skin mass was higher and concentrations of anthocyanin in the skin and tannin in the skin and seed were lower in 2006 than in 2004. In 2006, more frequent sampling revealed that the high ABA-GE levels had been sustained through maturation. There appears to be no association between levels of ABA-GE and berry phenolics.
After treating berries at veraison with ABA, levels of ABA found inside and on the surface of the pulp/skin were initially very high, as expected, and then rapidly declined. It is well known that ABA is photosensitive and isomerizes to a one-to-one mixture of trans-ABA and natural cis-ABA (Zaharia et al. 2005). The trans isomer is thought to be biologically inactive but does, in light, provide a source of ABA by isomerization. In this study the half life of ABA on the berry surface was less than one week (data not shown). Likely, it was rapidly converted to the inactive form on the surface of the berry, which was the principal cause of the rapid decline in ABA, as there was little change to the total concentration of ABA catabolites following the treatment. The increase in DPA indicates that a small amount of ABA was taken up by the berry and catabolized. Our findings are different from those of other researchers who performed a similar experiment spraying ABA on Merlot berries at veraison (Deytieux-Belleau et al. 2007), but there were differences in materials and sampling methods. Whereas we applied natural ABA, Deytieux-Belleau and colleagues applied the racemic form, which contains 50% unnatural (R)-ABA. This unnatural form is degraded more slowly in plants and can cause an increase in ABA biosynthesis (Lin et al. 2005). Other researchers have determined that (R)-ABA and trans-ABA are inactive in biochemical processes in ripening grape berries (Yu et al. 2006, Pan et al. 2005). These observations could explain why Deytieux-Belleau and colleagues observed the ABA concentration to peak at 10 days after application. Their treatment led to an earlier onset of ripening and anthocyanin production. We did not find that ripening was advanced by treating berries with ABA, based on soluble solids accumulation and acid degradation, but we did find elevated anthocyanins at berry maturity. The enhancement of anthocyanins was less than that observed in Flame Seedless (Peppi et al. 2006, Peppi and Fidelibus 2008), a color-poor variety, but greater than that observed in Cabernet Sauvignon (Gagné et al. 2006).
Conclusions
Repeated measurements of ABA and its catabolites over development provide a broader understanding of ABA biosynthesis and turnover than single or infrequent measurements of ABA alone. The ABA catabolite DPA was found in high quantities in early developing Merlot berries, suggesting that ABA is required early in berry development and then catabolized. The DPA concentration rapidly declines before the onset of veraison, at which point the concentration of ABA-GE begins to increase and ABA appears to be catabolized not by oxidation but by conjugation.
We found that ABA supplied to berries at veraison was rapidly degraded photochemically and had little effect on concentrations of ABA catabolites, except for a transitory rise in DPA. The subsequent small increase in anthocyanin accumulation indicates that either ABA or DPA enhances anthocyanin synthesis, but there was no effect on tannin levels. This study provides baseline data that will be used to assess the effects of stress and hormone treatments on the hormone content and fruit composition parameters in developing grape berries.
Footnotes
Acknowledgments: Financial support from the National Research Council of Canada’s Crops for Enhanced Human Health (CEHH) Cluster Initiative is gratefully acknowledged. This is NRCC article # 48425. Vincor Canada is gratefully acknowledged for hosting the field studies.
The authors thank Steve Ambrose, Elyse Busch, Vera Čekić, Patrycja Galka, Dang Van, and Irina Zaharia at PBI for advice and technical assistance and thank Steve Marsh at AAFC-Summerland for help with sampling.
- Received December 2008.
- Revision received March 2009.
- Accepted March 2009.
- Published online September 2009
- Copyright © 2009 by the American Society for Enology and Viticulture
Literature Cited
Vol 60 Issue 3








{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
More from this TOC section
Similar Articles
