


Abstract
Removing basal leaves 2 weeks postbloom significantly reduced powdery mildew severity on clusters in each year of a study in a New York Chardonnay vineyard. In contrast, removing leaves 5 weeks postbloom had no effect. The effect was not significantly different whether one leaf or two above and below each cluster was removed. Shoot density of vertical shoot-positioned (VSP) vines was lower than that of Umbrella-Kniffen vines and was associated with a significant reduction in disease development in one year of the study. When VSP-training was combined with early leaf removal in the absence of fungicide sprays, it reduced mean disease severity by 32% relative to untreated clusters on Umbrella-Kniffen-trained vines. However, there was no effect of training system in the second year of the study. In South Australia, doubling the volume of irrigation water supplied to vines that received a standard reduced deficit irrigation program resulted in two- and seven-fold increases in foliar powdery mildew severity in two consecutive seasons. Results highlight that viticultural practices targeted primarily at controlling vine growth and crop quantity and quality can also significantly affect the development of powdery mildew.
Grapevine management to optimize fruit quality and/or yield is a fundamental tenet of viticulture. Justification for one method over another depends on the cultivar grown, the prevailing viticultural practices, the desired market for the fruit, economic costs, geographic location, and vineyard microclimate. One method of manipulating the vine is the training system, a primary role of which is to optimize light interception and distribution and attendant yields and fruit quality (Reynolds and Vanden Heuvel 2009). Direct sunlight interception by fruit has been associated with improved fruit quality and is generally desirable to some degree in most vineyards (Smart 1985). For a given training system, basal leaf removal is an additional method of improving sunlight exposure of fruit. In addition to increasing direct sunlight exposure, removing leaves around fruit increases the evaporative potential within the fruit zone, lowering humidity and making the cluster microclimate less conducive for the development of fungal diseases (English et al. 1990), particularly botrytis bunch rot (Gubler et al. 1987, Zoecklein et al. 1992). Furthermore, sunlight exposure can increase surface temperature of grapes (Smart and Sinclair 1976) and other fruit (Schroeder 1965). The surface temperature of sunlight-exposed grape berries can be 5 to 15ºC warmer than ambient air temperature (Smart and Sinclair 1976), potentially altering basic components of berry physiology (Downey et al. 2006). Notably, these temperature studies have focused on the physiological effects on the fruit rather than any attendant consequence on disease development.
Powdery mildew (PM), caused by the fungus Erysiphe [Uncinula] necator (Schw.) Burr., is a disease of economic importance in viticultural regions throughout the world. Growth of the fungus is almost entirely epiphytic, and thus is persistently and directly exposed to various components of the environment that do not affect other pathogens embedded entirely within infected tissues. In Israel, reduced PM incidences were associated with a training system and row spacing that allowed more sunlight into the fruiting zone (Zahavi et al. 2001); similarly, it has been hypothesized that sunlight exposure within the fruit-zone creates a microclimate unfavorable for this disease (Chellemi and Marois 1992). Furthermore, vines grown under sunlight from which UV radiation was filtered had significantly more PM than those exposed to unfiltered sunlight (Keller et al. 2003). UV radiation has been shown to alter fungal populations on plant leaves (Moody et al. 2001) and generally increases fungal mortality (Rotem et al. 1985), including that of E. necator (Willocquet et al. 1996). Previous work has shown that sunlight exposure inhibits PM development through at least two interactive and synergistic mechanisms—exposure of the causal fungus to UV radiation and heating of sun-exposed tissues into a range unfavorable for fungal growth—and that disease development regularly differs within canopies of variable densities in a manner inversely proportional to the degree of admitted sunlight (Austin 2010, Austin et al. 2011).
Irrigation is a standard viticultural practice in many regions, particularly arid and semiarid climates, and has been proposed to be of growing significance in the face of global climate changes (Schultz and Stoll 2010). So-called deficit irrigation techniques have been applied both to improve water use efficiency within vineyards and to improve grape and wine quality (McCarthy 1997). Water stress has been correlated with increased grapevine leaf temperature (Möller et al. 2007) when stressed vines cease to transpire and evaporatively cool, causing the canopies to become warmer than those of unstressed vines. Consequently, increased canopy temperature is at times used as an indirect estimate of whole vine water status.
Irrigation has been shown to elevate levels of the PM on tobacco by promoting growth of susceptible tissue (Cole 1966) and on wheat by increasing mesoclimatic relative humidity (Sharma et al. 2004). However, there are no published reports of irrigation affecting the development of any PM through an effect on the temperature of host tissues. Nevertheless, increases in temperature above an optimum temperature can have significant detrimental effects on the development of grapevine PM (Delp 1954).
Viticultural practices can have pronounced effects on the development of specific diseases and, ideally for optimum production, these effects should be considered within the broad context of an integrated crop management system. The primary objective of this study was to determine whether two standard viticultural practices used to maximize fruit quality by increasing sunlight exposure within the fruit-zone (a relatively open training system and basal leaf removal around clusters) might also provide benefits with respect to improved management of grapevine PM. A secondary objective was to determine whether different irrigation regimes influence PM development through effects on canopy temperature.
Materials and Methods
Training system and leaf removal
The experiment was conducted in 2008 and 2009 in a vineyard of Vitis vinifera Chardonnay on 3309C rootstock with 2.75 m row spacing and 1.83 m vine spacing, planted in 2005 at the New York State Agricultural Experiment Station in Geneva. Vines were trained to two different systems, vertical shoot-positioned (VSP) or Umbrella-Kniffen (in which arching of the canes allows more buds per linear unit of row than in the VSP system) (Table 1). Three panels of four vines each were randomly assigned and trained to either the VSP or Umbrella-Kniffen system beginning in 2006. Vines in both training systems were balanced pruned each year using a 20 + 20 formula (20 buds retained on the vine plus 20 more for every 454 g of pruning weight). For all treatments, 10 clusters in each of three replicate four-vine panels (30 cluster total per treatment) were arbitrarily identified and inoculated with a suspension of Erysiphe necator conidia at 75% capfall. Cultures used for inoculum were sourced from field populations of E. necator collected from local vineyards and maintained throughout the year on 1- to 3-month-old Riesling seedlings in the greenhouse. During the growing season, conidia for inoculations were multiplied on 2-month-old Riesling seedlings maintained in a covered shadehouse. Inoculum was prepared in the field by agitating sporulating leaves in 50-mL falcon tubes with 0.005% Tween solution in dH20 and was applied at 1 mL per inoculated cluster using a small spray-paint apparatus (Preval, Yonkers, NY). Subsequent quantification in the lab with the aid of a hemocytometer revealed concentrations of 5 x 104 to 1 x 105 conidia/mL. Basal leaf removal was performed in both seasons either 2 or 5 weeks following 75% capfall. At each timing, leaves were removed by hand at the node opposite the inoculated cluster as well as one node above and below (light) or two nodes above and below (heavy). Leaves were not removed adjacent to control clusters. Ambient temperature and relative humidity within the canopy were measured with calibrated HOBO dataloggers (Onset, Pocasset, MA) at hourly intervals throughout the growing season. Fungal diseases on vines were managed with minimal sprays of mancozeb and a highly refined petroleum oil (Stylet Oil, JMS Flower Farms, Vero Beach, FL), chosen for, respectively, their relative lack of activity against E. necator and efficacy against this pathogen but with minimal residual and vapor activity. Fungicides were applied using an over-the-row, hooded-boom sprayer, and inoculated clusters were protected during each application by sealing them within individual plastic bags. The bags were removed immediately after sprays had dried.
Disease progress was monitored weekly throughout the season and final assessments were made postveraison on 5 Aug 2008 and 25 Jul 2009 because of the rapid degradation of heavily infected clusters and the development of botrytis bunch rot. Powdery mildew disease severity was assessed by visually rating individual clusters on a continuous 0 to 100% scale, based on the proportion of cluster tissue covered with the pathogen; all ratings were made by a single individual (C. Austin). Training system and basal leaf removal effects on cluster disease severity were analyzed for significance using GLM. Pair-wise comparisons of means for the effects of each leaf removal treatment in the model were made using Tukey–Kramer HSD. These and all other statistical analyses were conducted using JMP statistical software (SAS Institute, Cary, NC).
Irrigation effects
The potential effects of irrigation on PM development were investigated in an own-rooted Chardonnay vineyard at the South Australia Research and Development Institute in Nuriootpa (Barossa Valley), South Australia. This vineyard was planted in 1994 with 3.5-m row spacing and 2.25-m vine spacing. The experiment was conducted using two consecutive rows, each of 72 vines, pruned to two-bud spurs and ~40 buds per vine, grown on a three-wire single-curtain, non-shoot-positioned trellis. The vines were divided into three blocks, where a block consisted of six opposite four-vine panels in each of the two rows (12 panels in total). Blocks were then subdivided into six two-panel plots, half of which were randomly assigned to the standard reduced deficit irrigation (RDI) regime, provided through a single dripper line that supplied water to all vines, and the other half were assigned to twice that water volume, provided through the common dripper line plus an identical second line with emitters supplementing only those vines.
Pruning weights for vertical shoot-positioned (VSP) and Umbrella-Kniffen (UK) vines after the 2009 season. Vines were balance pruned using a 20 + 20 formula each season.
Vines were irrigated once a week for 12 hr at either 4 or 8 L/hr via one emitter per line per vine. Irrigations began 6 Dec 2006 and 13 Dec 2007, with a total of 11 and 12 irrigations for the two respective seasons. In both seasons, vines were unsprayed and natural PM infections were allowed to develop. Foliar disease progress was monitored weekly throughout each season, and final assessments were made on 25 Feb 2007 and 23 Feb 2008. Disease severity was assessed by visually rating the adaxial surface of 15 leaves per plot on a continuous 0 to 100% scale, based on the proportion of leaf tissue covered with the pathogen; all ratings were made by a single individual (C. Austin). These ratings were averaged for each of the three replicate blocks per treatment. Midday leaf water potentials to estimate vine water status (Chone et al. 2001) were determined for three leaves per plot, using a pressure bomb on the day after every third irrigation. Surface temperatures of 10 leaves per plot were measured at these same times using a noncontact Mikron infrared radiometer MI-N14 (LumaSense Technologies, Santa Clara, CA). Air temperature and relative humidity in the canopy were measured as described in the New York experiment above. Foliar disease severity ratings for irrigation treatments were compared using Student’s t-test.
Results
Training system and basal leaf removal
In 2008, there was a significant (p < 0.0001) effect of training system on PM development on clusters (Table 2), with mean disease severity levels for vines trained to the VSP system reduced by 24% relative to those trained to the Umbrella-Kniffen system when averaged across all leaf-removal treatments (Figure 1). Leaf removal also had a significant (p < 0.0001) effect, a result of the early but not the late treatments (Table 2). For example, disease severities were reduced by 16 and 18% relative to the control treatment on VSP-trained vines subjected to the light and heavy removal levels, respectively, when applied 2 weeks postbloom, whereas neither level had an effect when applied 5 weeks postbloom (Figure 1). Integrating the effects of training system and early leaf removal, final disease severities on VSP-trained vines subjected to light and heavy leaf removal 2 weeks postbloom were 31 and 33% lower, respectively, than on vines in the Umbrella-Kniffen system without leaf removal. There was no significant interaction (p = 0.61) between leaf removal and training system. There was no difference in air temperature or relative humidity measured within the canopies of vines in the two different trainings systems throughout either season (Austin 2011).
Statistical analyses for training system and basal leaf removal effects on powdery mildew severity on fruit.
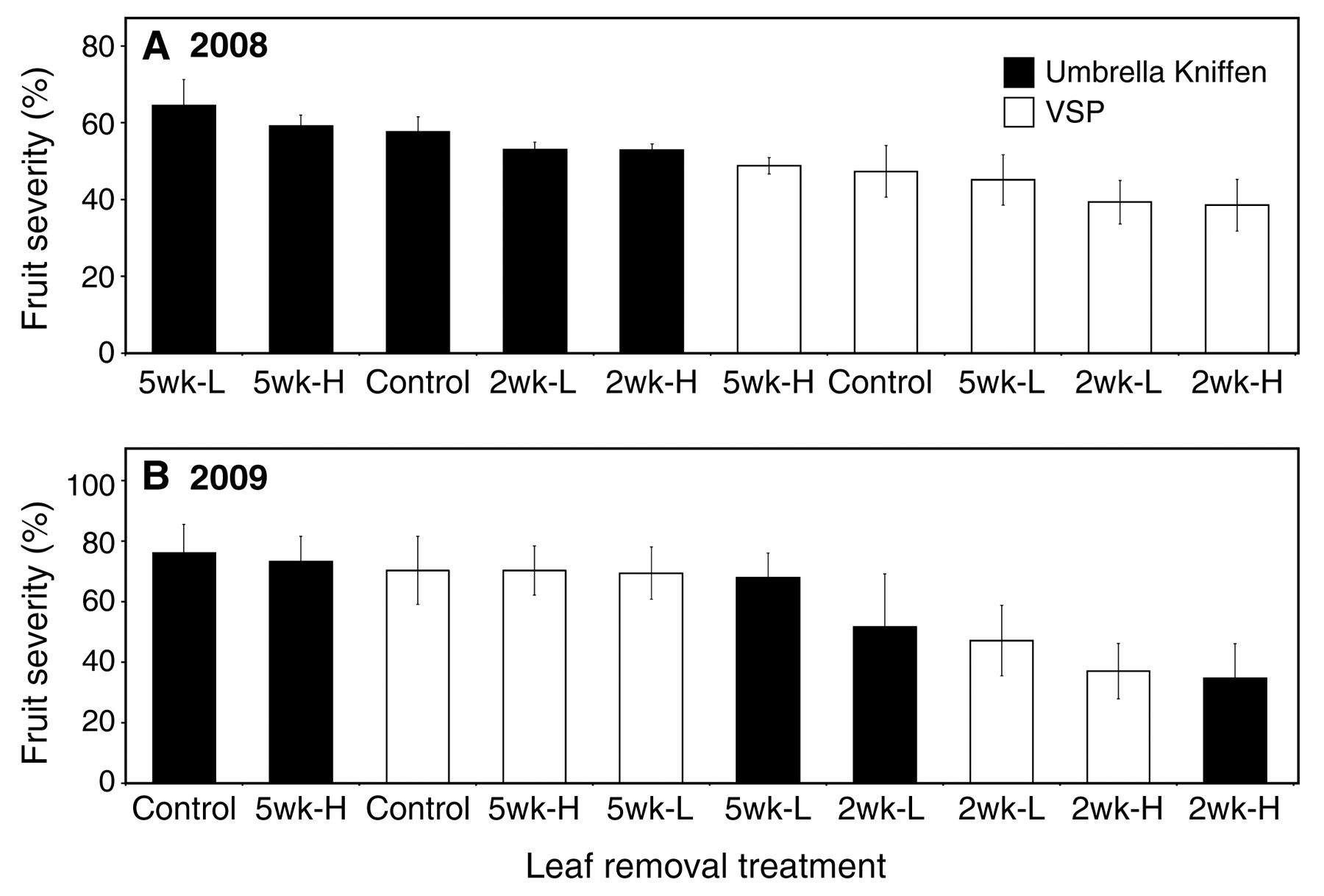
Powdery mildew severity on Chardonnay clusters in New York subjected to five different leaf-removal treatments in each of two vine-training systems for the 2008 (A) and 2009 (B) seasons. Clusters were inoculated at 75% capfall and fruit disease severity was visually assessed on a 0 to 100% scale on 1 Aug 2008 and 25 Jul 2009. Leaf removal was either 2 or 5 wk postbloom, and for each timing leaf removal was either heavy (H) or light (L) (two leaves or one leaf above and below each cluster, respectively). Each data bar represents the mean for 30 clusters per treatment, and error bars represent one standard error of the mean.
In 2009, when disease pressure was generally higher, training system no longer had an effect (p = 0.51). However, the effect of leaf removal was again significant (p < 0.0001), with an influence of treatment timing and a lack of an interaction between leaf removal and training system similar to those in 2008 (Table 2). As in the previous year, leaf removal treatments imposed 5 weeks postbloom had no effect on disease severity, whereas in contrast, the heavy removal imposed 2 weeks postbloom reduced severity by approximately one-half relative to the untreated vines in both training systems. Disease severity levels were somewhat higher in the light versus heavy removal treatment when both were applied early, although these differences were not statistically significant (p > 0.05) (Figure 1, Table 2).
Irrigation
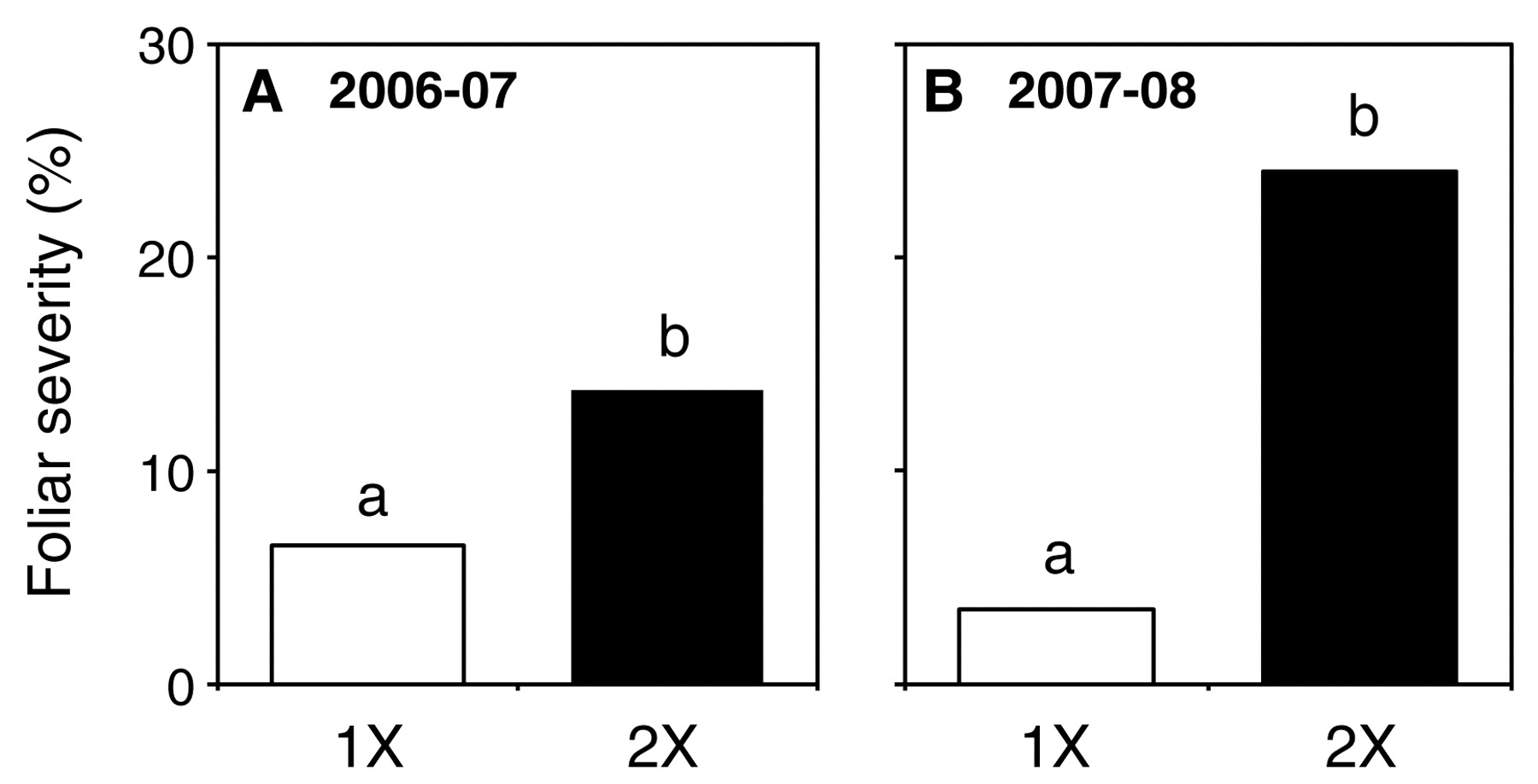
Vines that received double the standard RDI volume of water had significantly higher amounts of foliar PM than those receiving that standard regime, in both seasons (Figure 2). In 2007, disease severity on the doubly irrigated vines was twice that found on their standard RDI counterparts, whereas it was almost seven-fold greater in 2008. Midday measurements of leaf water potential and leaf surface temperature did not detect any significant differences in these values between the two irrigation treatments (Austin 2010). There were no differences in ambient temperature or relative humidity measured within canopies of the two treatments during either season (Austin 2010).
Discussion
Canopy management practices designed to reduce shading and promote the penetration of sunlight into the fruiting zone help limit the development of grapevine PM. Among the most basic decisions with respect to canopy management is the choice of training system. Although training systems are typically designed or adopted to optimize light interception for reasons of fruit yield and quality (Reynolds and Vanden Heuvel 2009), we hypothesized that a system chosen for these reasons may also be useful within an integrated approach to managing PM.

Foliar severity of powdery mildew on leaves of Chardonnay vines in Barossa Valley, S.A., irrigated with one (1x) or two (2x) dripper irrigation tubes for the 2006–2007 (A) and 2007–2008 (B) seasons. Vines were irrigated once a week for 12 hr at 4 L/hr per dripper. The vines were unsprayed and natural infection was allowed to occur. Means represent average disease severity from 15 leaves for each of three reps. Disease was assessed on adaxial leaf surfaces four weeks preharvest. Within panel A and B, bars not labeled with a common letter are significantly different (p < 0.001).
In the Finger Lakes region of New York (near Geneva), older V. vinifera plantings are often trained to an Umbrella-Kniffen system, although most newer plantings use systems, including VSP, that provide lower-density canopies. Thus, we chose these two training systems for comparison in the present study, and consistent with the above hypothesis, disease severity was one-fourth lower in 2008 for the VSP system, in which vegetative growth was also less relative to the Umbrella-Kniffen vines. Another study found similarly lower levels of PM in a sprawl or “free-positioned” training system than in a VSP system, in which VSP provided the more tightly compressed shoots, thereby providing the denser canopy structure (Zahavi et al. 2001). However, there was no difference in disease severity between the two training systems in our 2009 experiment, a season during which PM pressure was substantially greater throughout the region than in the previous year. For example, in an unrelated experiment using other Chardonnay vines near our experimental rows, cluster disease severity on unsprayed and uninoculated control vines was 99% in 2009 versus only 26% in 2008 (Wilcox and Riegel 2010a, 2010b).
A common canopy management technique for increasing sunlight exposure within the fruit zone is the removal of basal leaves near the clusters after fruit set. Although this practice was developed primarily to improve fruit quality characteristics (Reynolds et al. 1996, Smart 1985), disease management benefits also have been noted, particularly with botrytis bunch rot (English et al. 1990, Gubler et al. 1987), but also, in one report, with powdery mildew (Chelemi and Marois 1992).
In both years of our study, leaf removal 2 weeks after bloom (i.e., shortly after fruit set) significantly reduced disease severity compared to the control treatment, whereas leaf removal 5 weeks after bloom had no effect. Although grape berries are highly susceptible to infection by E. necator for the first 2 weeks after bloom, they quickly obtain age-related resistance thereafter and become virtually immune to new infection by 5 weeks postbloom (Gadoury et al. 2003). Thus, the detrimental effects exerted upon the fungus by sunlight via UV irradiation and/or the heating of exposed host tissues (Austin 2010), or through other mechanisms such as increasing host resistance to infection (Keller et al. 2003), may limit disease development more strongly when exposure occurs while berries are still young and highly susceptible. A significant reduction in PM development was noted in a study in which basal leaves were removed 3 weeks after bloom in one season, but unlike our results, there was a significant effect when leaves were removed 6 weeks after bloom in a different season (Chellemi and Marois 1992). However, it is difficult to determine any potential effect of treatment timing in that study, since the two timings were not directly compared in a common experiment and disease pressure in the experiment examining the 6-week postbloom timing was very low (15% severity on unsprayed controls), potentially confounding the results. It also is possible that the influence of timing on basal leaf removal effects, like the value of the procedure itself, may vary depending on location and weather (Guidoni et al. 2008). The authors speculated that their observed effect of leaf removal was due to the independent influences of improved fungicide coverage and creation of a microclimate less favorable for the disease (Chellemi and Marois 1992), anecdotally noting that disease was most prevalent on the interior portions of clusters least exposed to the sun, consistent with our data presented elsewhere (Austin 2010, Austin et al. 2011). And although the particular vines used in the present experiment were not examined for spray coverage, results from another experiment conducted in a different portion of this same New York vineyard showed that removing one leaf layer between a cluster and the outer edge of the canopy essentially doubled the quantity of spray deposited on that cluster (Austin et al. 2011).
Disease assessment data from the irrigation study in the Barossa Valley, South Australia, where doubling the irrigation volume led to a two- and seven-fold increase in foliar disease severity in two consecutive seasons, were consistent with the hypothesis that increased levels of irrigation in a hot, dry climate will increase PM severity by maintaining leaf temperatures in a range more conducive to the pathogen, through increased evapotranspirative cooling. However, data to support this mechanistic explanation for the observed phenomenon were lacking, as there were no differences measured in midday leaf water potentials or midday surface temperatures between irrigation treatments. The lack of measureable differences may be because there were no differences throughout the season or because differences were transitory and not present at the time that measurements were made. Vines assessed at midday in this location, independent of irrigation regime, were below a point of critical leaf water potential (less than −0.16 MPa) (Liu et al. 1978); thus, complete stomatal closure on leaves of all vines should have resulted in relatively uniform leaf water potentials (Patakas et al. 2005) and leaf temperatures as transpiration ceased. However, differential canopy surface temperatures can develop on Shiraz vines under variable irrigation regimes in Australia, with those vines that received more water remaining relatively cooler (Loveys 2005).
Irrigation can have numerous numerous effects on grapevine physiology (Williams et al. 1994), and it is possible that the increased foliar PM severity in response to increased irrigation resulted from factors in lieu of or in addition to any effects on leaf temperature. Furthermore, whereas our instruments indicated no difference in humidity within the canopy mesoclimate of vines subjected to the two irrigation regimes, current technology does not readily allow measures of the microclimatic zone immediately surrounding leaves and berries. The direct relationship between quantities of atmospheric water vapor and the severity of powdery mildew on grape foliage has been documented, but humidity in the leaf boundary layer zone inhabited by the fungus was likely higher than that measured in the bulk air around it (Carroll and Wilcox 2003). Therefore, it is also possible that increased irrigation elevated disease severity by increasing the humidity of the microclimate within which the pathogen was functioning, although we were unable to detect it. Regardless of the underlying mechanism involved, our results underscore that viticultural practices targeted primarily at general vine growth, yield, and fruit quality can also significantly affect the development of powdery mildew. This concept, and these particulars, should be considered when devising integrated pest management and integrated crop management programs.
Conclusion
Viticultural practices that alter sunlight distribution within the fruit zone also alter the development potential of PM on fruit, with disease severity reduced in systems providing increased sunlight exposure. In a New York vineyard, an Umbrella-Kniffen training system was at higher risk for PM development than a VSP system, which had a lower shoot density; however, seasonal climatic variation apparently overrode this effect of training system on disease development in one of the two seasons studied. Basal leaf removal consistently reduced disease severity on fruit within both training systems when performed 2 weeks postbloom, while berries were still highly susceptible to infection. In contrast, leaf removal 5 weeks postbloom, when berries were no longer susceptible to new infections, had no effect on disease severity. Doubling irrigation water volume supplied to vines in the dry Barossa Valley region in South Australia increased foliar disease severity by a factor of two to seven. Our limited measurements of midday leaf water potentials and leaf surface temperatures could not mechanistically explain the phenomenon; however, the consistent results obtained suggest an effect of irrigation on pathogen development either directly though altering the canopy microclimate or indirectly through a physiological response of the grapevine. These results underscore the attendant consequences that viticultural practices targeted primarily at manipulating vine growth and crop quantity and quality can also have on the development of powdery mildew.
Acknowledgments
Acknowledgments: This work was supported in part by grants from the USDA Viticulture Consortium–East, the Lake Erie Regional Grape Program, the Grape Production Research Fund, and the New York Wine and Grape Foundation.
Footnotes
-
The authors gratefully thank Trevor Wicks and Michael McCarthy of the South Australian Research and Development Institute for their technical advice and assistance in our Australian trials.
- Received August 1, 2010.
- Revision received November 1, 2010.
- Accepted November 1, 2010.
- Published online December 1969
- © 2011 by the American Society for Enology and Viticulture
Literature Cited
Vol 62 Issue 2








{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
More from this TOC section
Similar Articles
