


Abstract
Water and nutrients exist together in close association because plant-available nutrient ions are dissolved in the soil solution and nutrient uptake by grapevines depends on water flow through the soil-root-shoot pathway. Leaf transpiration generates the tension necessary for the roots to absorb this essential solution, but in a drying soil, uptake of water and nutrients becomes progressively more difficult for grapevines. In addition, application of nitrogen fertilizer can increase the vine’s susceptibility to drought, because nitrogen favors shoot growth over root growth. However, because growth is more sensitive than photosynthesis to both water and nitrogen shortage, deficit irrigation may be used in conjunction with limited nitrogen application to control canopy development, yield, and fruit composition. Growth is the “pacemaker” for nutrient uptake by the vine, hence the growth reduction induced by water deficit also decreases vine nutrient requirements. Nevertheless, reducing water or nitrogen supply can be perceived as a stress by the vine, and its response depends on developmental status. For instance, water deficit applied before fruit set may reduce cluster and berry numbers, especially if combined with nitrogen shortage. Properly regulated deficit irrigation in combination with low to moderate rates of nitrogen application between bloom and veraison reduces canopy size, berry size, and yield, accelerates ripening, improves fruit color, and reduces disease incidence. However, this strategy also reduces yeast-assimilable nitrogen in the fruit, thereby increasing the risk of sluggish or stuck fermentation. Moreover, if the water or nitrogen deficit becomes too severe, fruit quality suffers from both limited assimilate supply and excessive fruit exposure to sunlight. The relationship between vine nutrition and deficit irrigation clearly requires careful guidance to make it a happy one.
- Vitis vinifera
- water stress
- irrigation
- nutrient uptake
- nitrogen
- photosynthesis
- transpiration
- growth
- yield components
- fruit composition
- PRD, partial rootzone drying
- RDI, regulated deficit irrigation
- rh, hydraulic resistance
- rs, stomatal resistance
- RWC, relative water content
- Δ, gradient
- Ψ, water potential
Water and nutrients are basic requirements for plant growth and performance, and leaf water supply constitutes the overwhelming limitation on plant productivity (Kramer and Boyer 1995). Under natural conditions, water is supplied by snow and rainfall and temporarily stored in the soil for extraction by plant roots. However, the amount of rainfall varies greatly from region to region and from season to season, which may drastically impair vine performance and the economics of viticulture in some regions and some years. But water availability depends not only on how much rainfall a vineyard receives but also on when the rain falls and how rapidly it evaporates. In addition, soil water-holding capacity and hence the amount of plant-available water varies with soil depth, texture, and organic matter content. Variation in soil moisture due to differences in water holding capacity and effective rootzone has a pronounced impact on vine performance both between and within vineyards (Hall et al. 2002, Lamb et al. 2004).
Soils vary not only in their capacity to store water, but also in the amount and composition of mineral nutrients they contain and in the extent to which these nutrients are available for uptake by the roots. Nutrient storage capacity and accessibility are influenced by soil texture, rooting depth, and organic matter content, but availability is modified by soil moisture and pH. As both oversupply and shortage of water or nutrients result in unbalanced vine growth and poor production, irrigation and nutrient application (from organic or inorganic sources) are powerful tools for improving vine performance. They allow us to manipulate environmental variables when properly integrated in a soil management program. They can be used as supplements to compensate for shortcomings of climates and soils: that is, to provide adequate moisture and nutrients when nature does not supply them during the critical stages of the seasonal growth cycle. Thus the degree of dependence on irrigation and fertilizer application varies with climate and growing region. Of course, while irrigation could also be used to leach overabundant nutrient ions from the soil profile, it can do nothing to remove excessive soil moisture. That problem is best addressed by using proper site selection and other soil management options, such as cover crops or permanent swards. In addition, higher planting density or lighter pruning could be used to increase water use early in the season due to the earlier canopy development.
Abundant water and nutrient availability is a problem that mainly concerns cool (and some tropical) growing regions that experience frequent summer rainfall. However, the majority of the world’s grapes are grown under Mediterranean climatic conditions, which experience cool, moist winters and warm, dry summers. Following a wet start with a water-saturated soil profile, the soil progressively dries down, and the growing season is often characterized by low soil moisture and high vapor pressure deficit (in other words, warm or hot and dry air). Many vineyards are to varying degrees dependent on irrigation. Decreasing availability of irrigation water in California and Australia and increasing frequency of summer drought associated with global warming in Europe (Schultz 2000) in combination with the recognition that water availability influences canopy development, vine microclimate, yield, and fruit composition, have led to widespread adoption of deficit irrigation strategies. Increasingly, consumers and legislators are demanding sustainable production practices with a concomitant reduction in vineyard input and environmental impact, putting pressure on the use of fertilizers. The increasing incidence in Europe and North America of a phenomenon termed atypical aging has been attributed to reduced use of nitrogen fertilizer and increased use of permanent swards or vineyard floor covers, particularly in dry seasons (Löhnertz et al. 2000). Affected white wines develop an off-flavor reminiscent of acacia flower, naphthalene, or floor polish at the expense of varietal flavor. However, despite their obvious importance, we know surprisingly little about the interactions of water and nutrient availability and their influence on vine function and productivity.
Of all mineral nutrients, nitrogen (N) is the one that grapevines require in greatest quantity, that most often limits growth, and that is the most potent in terms of influencing fruit quality, making it the single most important mineral nutrient. When grapes are harvested, some of the N fixed in the fruit is permanently removed from the soil. This loss amounts to 1.0 to 3.0 kg/t of fruit removed (Williams and Matthews 1990, Conradie 2005), depending on the N concentration of the juice and whether or not stalks and pomace are recycled to the vineyard, and has to be replaced by addition of fertilizer or by biological N fixation using leguminous cover crops. Nitrogen is a structural component of a range of critically important plant constituents. Nucleic acids contain it and in turn make up the genetic information contained in the vine’s DNA. Nitrogen is a key component of amino acids which, linked together, make up the structural proteins and enzymes that drive all biochemical reactions. It also is an integral part of chlorophyll, responsible for intercepting and capturing sunlight in plants, of hormones used for communication between different plant organs, and of certain secondary metabolites, some of which also contribute to wine flavor. Of course, nutrients other than N also have important functions in the vine. However, a review on potassium (K) nutrition in grapes has recently been published (Mpelasoka et al. 2003), and uptake and partitioning of a range of nutrients in addition to N is discussed by Conradie (2005). Uptake of phosphorus (P) and several other nutrients depends largely on symbiotic mycorrhizal fungi, a topic covered by Schreiner (2005). Therefore, this review will focus primarily on physiological aspects of N nutrition and how it interacts with water deficit, while briefly touching on one other macronutrient (P) and one micronutrient (boron, B). I will draw heavily on conclusions drawn from investigations with other plant species. I will also discuss implications for yield formation and fruit quality as well as some practical implications for vineyard water and nutrient management.
Physiology of Water and Nutrient Uptake and Transport
During the transition from dormancy to active growth of a grapevine in early spring, water uptake by the roots and transport to the shoots are driven by positive root pressure (Alleweldt 1965, Sperry et al. 1987) induced by remobilization of stored nutrients and starch and unloading of osmotically active solutes into the xylem. Root pressure serves to dissolve and push out air bubbles that have formed during the winter in the dehydrated xylem. In addition to restoring vascular function, it may also be necessary to rehydrate dormant buds to enable them to resume growth and break. However, soon after budbreak transpiration from the expanding leaves produces a negative pressure (tension), which maintains the upward flow of water in the xylem. Therefore, water flows from the soil toward the roots because of suction at the root surface, which is generated osmotically (by root pressure) or hydrostatically (by transpiration). During most of the season, transpiration is the main driving force for water uptake and movement in the xylem up the vine to the leaves against gravity. Water flow from the soil to the root, from the roots to the leaves, and from the leaves to the atmosphere is caused by a water potential gradient (ΔΨ). In analogy to Ohm’s law, the flow (F) of water from the soil to the leaves can be described by the equation F = ΔΨ rh−1, where ΔΨ = Ψsoil – Ψleaf. The parameter rh describes the resistance to water flow due to friction between water and conduit walls and between the H2O molecules themselves. Most of the rh is imposed by the vine’s hydraulic architecture (plumbing layout), which is mainly determined by the shape, size, and arrangement of xylem conduits (water pipes), the permeability of the pits connecting individual conduits, as well as the total length of the flow pathway and the number and shape of bends (vine size and shape). The rh decreases with rising temperature, and transport of nutrient ions (especially cations such as K+) reduces rh by increasing pit permeability (Zwieniecki et al. 2001). In other words, a higher nutrient concentration in the xylem sap increases the sap flow rate and, conversely, nutrient deficiency strongly reduces sap flow even under well-watered conditions.
Water will flow from the soil to the vine as long as Ψxylem is lower than Ψsoil , and from the xylem to the leaf mesophyll as long as Ψleaf is lower than Ψxylem. The Ψleaf of well-watered vines fluctuates during the day following both opening and closing of stomata at dawn and dusk, and evaporative demand (vapor pressure deficit) of the air. Transpiration-driven water flow is minimal at night (although there is still water flow to satisfy cuticular transpiration, growth requirements, and phloem counterflow), so that Ψleaf is at its daily maximum (Schultz and Matthews 1988, Boyer et al. 1997). In other words, soil water status determines the baseline Ψleaf in the near absence of transpiration (Tardieu and Simonneau 1998). However, extraction of water by the roots and transport to the shoots becomes increasingly difficult as the soil dries. To maintain the ΔΨ driving water transport, Ψleaf must decrease, so predawn Ψleaf can be used as an indicator of Ψsoil. Transpiration during the day decreases Ψleaf below the pre-dawn level, and Ψleaf at a particular time of day is the result of both Ψsoil and transpiration linked to evaporative demand (Smart 1974, Tardieu and Simonneau 1998). As the soil dries the resistance to water flow will eventually become so great that the vine can no longer maintain a sufficient ΔΨ to sustain transpiration. The resulting imbalance between water uptake by the roots and water loss from the leaves can induce drought stress and leaf wilting. In his influential review, Hsiao (1973) defined mild plant water stress as a decrease of Ψleaf by several bars or of relative water content (RWC, percentage of tissue water relative to the water content of the same tissue at full turgor) by 8 to 10% below corresponding values in well-watered plants under mild evaporative demand. Moderate water stress was defined as a decrease of Ψleaf by more than several bars but less than 1.2 to 1.5 MPa or of RWC by 10 to 20%, and severe water stress as lowering of Ψleaf by >1.5 MPa or of RWC by >20%.
For xylem sap to sustain the tension required to pull water from the soil, the plumbing system has to be airtight. However, the water column inside the xylem can break if it is under too much tension (such as excessive transpiration due to wind or heat combined with dry soil) or if the tension is suddenly relieved (such as by shoot, leaf or cluster removal, or physical impact). This phenomenon is termed “cavitation,” in which the xylem vessels fill with H2O vapor or air bubbles. Gas blockages (embolisms) greatly increase rh (to infinity in the affected vessel itself), which renders the vessels nonfunctional and can lead to canopy desiccation (Schultz and Matthews 1988). For the gases to dissolve and conduits to refill with liquid water, the xylem pressure must rise to near atmospheric or above (Sperry et al. 2002). If cavitation occurs in the trunk, repair normally requires positive root pressure (during rainfall or humid nights leading to guttation) or starch remobilization and loading of sugar into the vessels. Luckily, the trunk and shoots are less prone to cavitation than the minor veins in the leaves (Nardini et al. 2001), where the high solute concentration may repair embolisms very rapidly (De Boer and Volkov 2003, Trifilò et al. 2003). However, root xylem is much more vulnerable to cavitation than shoot xylem (Sperry et al. 2002), which may be a problem with shallow root systems, because the soil dries from the top down. Shoot damage can be avoided if deeper roots are present, since cavitation in the surface roots will shift water uptake down to wetter soil layers (Sperry et al. 2002). Moreover, grapevines growing under conditions of moderate water stress develop narrower xylem vessels than vines growing with abundant water supply (Lovisolo and Schubert 1998). The smaller vessel diameter increases rh, which leads to reduced water loss and increased cavitation resistance.
Water uptake is proportional to root surface area. Because of their often dense root system in the topsoil, grapevines can remove water effectively from the surface soil layers. Extracting water from lower soil layers is more difficult, so that the surface soil dries more quickly than the subsoil. Under nonirrigated conditions roots continue to grow into deeper, wetter soil layers, whereas the roots of irrigated plants proliferate mostly in the topsoil (Hsiao and Xu 2000). Moreover, although roots can extract water more easily from water-saturated sand (owing to the large pore sizes) than loam, a coarse soil dries much more rapidly, so that water uptake becomes much more difficult in the sand as Ψsoil decreases. Since plant-available nutrient ions are dissolved in the soil solution, nutrient uptake also depends on water flow through the soil-root-shoot pathway. Nutrients are often concentrated in the biologically active surface soil, but water and nutrient availability varies greatly in both space and time. To complicate matters further, different nutrients are often available in different locations; nitrate (NO3−) leaches into the subsoil much more rapidly than potassium (K+) which diffuses much faster than phosphate (H2PO4−). As a consequence, superficial roots may take up soil-immobile nutrients (such as K and P), while deeper roots procure soil-mobile nutrients (such as NO3−).
Water entering the root initially moves through the root’s epidermis and cortex tissue along both symplastic (intracellular) and apoplastic (extracellular, including cell walls) routes (Steudle and Peterson 1998). Because there are no membranes along the apoplast, hydraulic water flow dominates along this path, whereas symplastic movement is influenced by osmotic gradients. Rapid transpiration favors apoplastic water flow and thus reduces rh, but when there is little or no transpiration (at night or with dry soil), rh increases substantially, because water flow is driven by osmotic gradients (across membranes) (Steudle and Peterson 1998, Steudle 2000). Rather than moving exclusively in one or the other compartment, a third possibility for water flow is straight across cell walls, membranes, and through cells (transcellular pathway). The endodermis that separates the cortex from the stele has thickened radial and transverse cell walls called Casparian strips (or Casparian band), which are impregnated with suberin and lignin. This waterproofing hydraulically separates the cortical and stelar apoplasts and forces water and its nutrient cargo to pass across the cell membranes into the symplast. Unlike cell walls, cell membranes are semipermeable; they act like selective sieves, permitting entry to some ions and not to others. Membrane pores created by special channel and transport proteins regulate the passage of water and nutrient ions. Many channels, especially in the endodermis and stele, are specific for water (aquaporins) and control water flow by opening (decreasing rh) or closing (Steudle 2000); others are specific for cations or for particular ions (ion channels); still others are specific for neutral solutes. Ions can move passively by simply diffusing across channels down an electrochemical potential gradient (usually from high to low concentration). In contrast, ions moving actively (such as NO3 ,− H2PO4−, or K+) are “pumped” through transport proteins or carriers against their electrochemical potential (usually from low to high concentration), which requires an input of energy in the form of adenosine-5′-triphosphate (ATP) (Grossman and Takahashi 2001). Active transport enables grapevines to concentrate nutrient ions inside the roots well above their concentration in the surrounding soil solution (Keller et al. 1995, 2001b), although some concentration is also possible due to electrical potential gradients. Some ions (NO3−, K+) are taken up passively across ion channels when their availability in the soil solution is high (generally in the mM range, for instance, after fertilizer application) and actively (and very selectively) across carriers, when availability is low (usually in the μM range) (Tester and Leigh 2001). This ensures that roots absorb nutrient ions over a wide range of external concentrations.
Ultimately, most water and ions return to the apoplast when the water is released into the root’s xylem conduits (Sattelmacher 2001). Since the cell walls of these conduits are waterproof, entry into the xylem is another bottleneck in the vine’s plumbing system after the passage through the endodermis. The presence of aquaporins and selective channels and carriers also controls access to the xylem (Tester and Leigh 2001, De Boer and Volkov 2003), which transports the water and its dissolved nutrients to the shoots and delivers them initially to the apoplastic compartment of transpiring organs. Grapevines lose most water by transpiration, but small amounts are used for cell expansion and metabolism and phloem transport. The water absorbed by the roots to balance this growth water and phloem counterflow (transpiration-independent water flow) is sufficient to transport nutrients, so that nutrient uptake and long-distance transport in the xylem are independent of transpiration (Tanner and Beevers 2001). Of course, the increase in water flow due to transpiration during the day results in a corresponding increase in solute delivery rate (Peuke et al. 2001). Thus rapid transpiration increases nutrient uptake, especially if nutrient availability in the soil is high (Hsiao 1973, Alleweldt et al. 1984).
Thanks to transpiration and phloem counterflow, xylem sap transports large amounts of nutrient ions in addition to organic molecules such as amino acids as N carriers (see below), organic acids such as malate and citrate as carriers of metal ions (iron, copper, zinc), and hormones such as abscisic acid (ABA) and cytokinins (Sattelmacher 2001). Solutes can also diffuse in the nonxylem apoplast, but this diffusion is orders of magnitude slower than the movement in the xylem. Long-distance transport in the xylem is by mass flow, and the flow rate increases exponentially (actually to the power of 4) with increasing diameter (decreasing rh) of the xylem vessels. Thus the large vessels formed in spring can transport water much more rapidly than the narrower vessels formed during summer at the cost of being more vulnerable to cavitation. Moreover, older vines with more annual rings and a larger trunk diameter are able to transport much more water to support a larger leaf area than can young vines. The roots and trunk also act as important water reservoirs, which are depleted in the morning, when the stomata open and transpiration rises, and replenished in the afternoon (Schultz and Matthews 1988, Steppe and Lemeur 2004). Older vines can store more water, and this buffering capacity could make them less vulnerable to xylem cavitation.
Not surprisingly, the major resistance to water flow through a grapevine is in the roots, although rh increases with increasing transport distance (for example, with increasing plant size) (Sperry et al. 2002). The other main resistance is in the leaves, at the terminal component of the transpiration stream, where water flows through orders of veins in series and in parallel, leaves the xylem network, and moves into and around the mesophyll cells, before it evaporates into the air spaces and diffuses out of the stomata during transpiration (Comstock 2002, Sack et al. 2003). Transfer of water and solutes from the xylem into the leaf mesophyll (i.e., back into the symplast) again requires aquaporins and energy-dependent carriers.
A Case Study in Nutrition: Nitrogen Uptake and Utilization
Owing to the rapid nitrification rates in most aerobic soils, nitrate (NO3−) is the primary source of N for grapevines (Keller et al. 1995, 2001b), although vines are also capable of taking up ammonium and, almost certainly, amino acids (Fischer et al. 1998, Grossman and Takahashi 2001). The concentration of nitrate in the soil water is generally orders of magnitude lower than the concentration inside the plant (Crawford 1995, Keller et al. 1995, 2001b). Therefore, roots absorb nitrate actively by means of H+/NO3− cotransport using H+-ATPase embedded in the cell membrane (Crawford 1995). The micronutrient boron (B) is essential to keep the ATP pump going, so that B deficiency strongly reduces the ability of roots to absorb NO3− (Camacho-Cristóbal and González-Fontes 1999).
Once inside the root cells, nitrate can be moved to the vacuoles for temporary storage, loaded into the xylem for transport to the shoots, or assimilated into amino acids for metabolism and transport (Loulakakis and Roubelakis-Angelakis 2001, Tester and Leigh 2001, Figure 1⇓). A recent review (Loulakakis and Roubelakis-Angelakis 2001) has covered N assimilation in grapevines in detail, and the following only gives a very brief summary of the assimilation process. It proceeds in a series of steps using a total of at least 10 electrons provided by NADH, NADPH, or reduced ferredoxin (Fdred). The starting point is the reduction of nitrate to nitrite (NO2−) by nitrate reductase (NR: NO3− + 2H+ → NO2− + H2O). Nitrate reductase is stimulated by nitrate and cytokinins, but requires light (in the leaves) or carbohydrates (in the roots) for full activation, and is suppressed by amino acids (especially glutamine). Grapevines can distribute large amounts of nitrate or store it for later use in the cell vacuoles without deleterious effects. In contrast, nitrite is toxic and is rapidly reduced to ammonium (NH4+) by nitrite reductase (NiR: NO2− + 8H+ → NH4+ + 2 H2O). In the roots, nitrate is sufficient to activate NiR, whereas in the leaves light is also required. Like nitrite, ammonium is toxic to plants and is usually rapidly incorporated into amino acids. Ammonium assimilation is normally catalyzed by glutamine synthetase (GS) and glutamate synthase, also known as glutamine-2-oxoglutarate aminotransferase (GOGAT), in two sequential reactions: glutamate + NH4+ → glutamine; and glutamine + 2-oxoglutarate → 2 glutamate. The first step requires energy provided by ATP and a divalent cation (Mg2+, Mn2+, or Co2+) as cofactor. Glutamine produced by GS stimulates GOGAT activity producing two molecules of glutamate. Both GS and GOGAT are activated by light and sucrose, and inhibited by amino acids. One of the two glutamate molecules is used to regenerate the GS/GOGAT cycle, the other to supply amino acids for metabolism. In the roots, glutamate can also be converted back to glutamine (by a slightly different form of GS) for export in the xylem to the shoots. In an alternative pathway for NH4+ assimilation, glutamate dehydrogenase (GDH) catalyzes a reversible reaction that can either form or degrade glutamate (2-oxoglutarate + NH4+ ↔ glutamate + H2O). As a stress-related enzyme GDH assimilates ammonium in tissues with excessive ammonium concentration, thus complementing the GS/GOGAT cycle. However, GDH can operate in the reverse direction, oxidizing glutamate when carbohydrates are depleted. Under conditions of limited photosynthesis, GDH can help remobilize proteins and degrade amino acids to supply carbon skeletons for continued ATP regeneration.

Simplified diagram of nitrogen uptake, assimilation, and distribution in grapevines.
Nitrogen uptake and assimilation require large amounts of energy to convert stable, low-energy inorganic compounds from a dilute soil solution into high-energy organic compounds concentrated inside a grapevine. In addition, the production of amino acids also requires a supply of carbon skeletons. Nitrate acts as a signal that induces the vine to activate N assimilation and divert carbon away from starch production to manufacture of amino acids and organic acids such as malate (Stitt 1999). Therefore, N assimilation proceeds rapidly in vines with high carbohydrate status but slows down (leading to tissue nitrate accumulation) as photosynthetic sugar supply decreases (Perez and Kliewer 1982). Assimilation is cheaper in leaves, which can use spare photosynthetic energy to assimilate N, than in roots where it depends entirely on phloem-imported sucrose. The relative contribution of roots and leaves to N assimilation depends on a number of factors, including variety and rootstock, and the amount of nitrate absorbed by the roots (Alleweldt and Merkt 1992, Keller et al. 1995, 2001b). Since nitrate cannot be transported in the phloem, it has to be assimilated once the xylem has delivered it to the leaves, or stored in vacuoles. When carbohydrate status is high and soil nitrate supply low, N is readily assimilated in the roots and exported as glutamine in the xylem (Alleweldt and Merkt 1992, Keller et al. 1995, 2001b, Llorens et al. 2002). However, because of the limited capacity of the roots, the proportion of N assimilated in the leaves increases with increasing N uptake (Keller et al. 1995, 2001b, Zerihun and Treeby 2002). To reduce luxury consumption, glutamine is exported from the leaves in the phloem as a signal for the roots to limit nitrate uptake (Gessler et al. 1998, Lemaire and Millard 1999). This feedback regulation normally coordinates uptake with the vine’s demand. However, when the nitrate concentration in the soil water is high (such as after fertilizer application or incorporation of cover crops), uptake can exceed the capacity for assimilation (Perez and Kliewer 1982), especially under conditions favoring rapid transpiration (warm, sunny days, and high soil moisture), and nitrate may accumulate in the leaves (Scheible et al. 1997, Zerihun and Treeby 2002). High leaf-nitrate reduces the amount of sucrose available for export, depleting root starch reserves and inhibiting root growth, which strongly decreases the root:shoot ratio (Keller et al. 1995, Keller and Koblet 1995, Scheible et al. 1997). That has important implications during the establishment phase of vineyards. Heavily irrigating and fertilizing young vines in order to harvest a crop in the year after planting may be detrimental to vine performance in the long term.
A large portion of the glutamine and nitrate continuously arriving in the leaves is redistributed as glutamine via the phloem (Figure 1⇑) to the various sink organs for use in growth and metabolism. Glutamine and glutamate can also be converted to other amino acids for incorporation into proteins or storage. When conditions favor high rates of both C and N assimilation, surplus glutamine is converted to arginine, which is thought to be the major N storage compound in grapevines (Kliewer 1967, Schaller et al. 1989). However, accumulation of storage reserves only occurs when all other plant requirements (such as growth and fruit ripening) have been satisfied, that is, when supply of resources exceeds demand (Lemaire and Millard 1999). In addition, nitrogenous compounds (as well as mineral ions) are remobilized from shaded leaves in the interior of dense canopies and recycled to the shoot tips for production of new leaves with better sun exposure. Remobilization also occurs in senescing leaves at the end of the season for export to the perennial parts of the vine for storage (Loulakakis et al. 2002, Conradie 2005). Stored N reserves can buffer temporary shortages in supply and support new growth in spring. Because of the rapid shoot growth early in the season, a vine’s nutrient demand is greatest between budbreak and bloom, even though most of the N uptake from the soil occurs after bloom, provided there is sufficient soil moisture (Peacock et al. 1989, Conradie 2005, Schreiner 2005). Limited availability of N reserves due to inadequate refilling in the previous growing season can restrict early shoot growth and canopy development (Keller and Koblet 1995). Storage reserves reach a minimum around bloom (Löhnertz 1991), making vines vulnerable to deficiency if insufficient N is available in the soil.
Limiting Supply: Responses to Water and Nutrient Deficits
Grapevines sense water deficit hydraulically and biochemically.
The water balance of a grapevine is determined by the amount of water lost in transpiration to the atmosphere and the amount of water absorbed from the soil. A vine can become water stressed as a result of both decreased Ψsoil, which generally occurs progressively over a period of time, and fluctuating transpiration rate, which occurs with daily and seasonal changes in vapor pressure deficit. To avoid xylem cavitation, the flow of water from the soil to the leaves must balance the water lost through the stomata, which can be written as E = (Ψleaf – Ψair) (rs + rb)−1 = (Ψsoil – Ψleaf) rh−1. For a given microclimate (determining Ψair) and soil water status (Ψsoil), the transpiration rate (E) is determined by the stomatal resistance (rs) and boundary layer resistance (rb) of the leaves, whereas rh determines Ψleaf at that transpiration rate (Tyree and Zimmermann 2002). Thus rh defines how wide the stomata can be open without desiccating the leaves, and the major function of stomata may be to avoid damaging water deficits causing xylem cavitation (Jones 1998, Brodribb and Holbrook 2003). Grapevines are quite susceptible to cavitation and are usually regarded as isohydric species (Düring 1987) whose sensitive stomata rapidly increase rs and decrease transpiration in response to low Ψsoil, which enables them to maintain almost constant Ψleaf throughout the day and regardless of Ψsoil. This effect can override the influence of high light intensity on stomatal opening, so that soil water deficit often leads to a midday increase in rs in the vineyard (Loveys and Düring 1984, Correia et al. 1990, 1995), although varieties that are less susceptible to xylem cavitation (such as Syrah) require less sensitive stomata (in other words, maintain lower rs and higher transpiration rates) and markedly decrease Ψleaf during the day and in response to drought (Escalona et al. 1999, Schultz 2003). However, since the cuticle allows small amounts of water vapor to pass through, leaves cannot avoid water loss completely, no matter how tightly the stomata are closed (Boyer et al. 1997). This problem is particularly acute when drying wind reduces rb and stimulates transpirational water loss.
The hydraulic effect of Ψleaf on rs is amplified by abscisic acid (ABA) which is synthesized in dehydrated cells. In addition to the ABA produced in drying leaves, the amount arriving from the roots can increase substantially in response to reduced soil moisture (Bray 1997, Davies et al. 2002, Wilkinson and Davies 2002). As Ψsoil declines, the roots produce increasing amounts of ABA, which is transported in the xylem to the leaf guard cells, where it triggers stomatal closure. This response to ABA is independent of Ψleaf and vapor pressure deficit, but is related to an increase in xylem sap pH, which enhances ABA delivery to the guard cells (Bacon et al. 1998, Wilkinson and Davies 2002). Even when only a portion of the roots experience dry soil, the ABA produced by that portion is sufficient to trigger stomatal closure, even when the remainder of the roots take up enough water to maintain high Ψleaf (Stoll et al. 2000, Comstock 2002, Lovisolo et al. 2002). In contrast to ABA, root-sourced cytokinins in the xylem sap decrease in response to soil drying (Davies et al. 2002, Yang et al. 2002). When rainfall or irrigation replenishes soil moisture, the roots stop producing ABA, and the ABA in the leaves is readily degraded, so that the stomata can reopen.
Shoot growth is extremely sensitive to water deficit. Roots must absorb more water than what is lost in transpiration to enable growth, since growth is mainly caused by cell expansion due to water import (Boyer 1985, Hsiao and Xu 2000). Water uptake by cells is driven by accumulation of solutes (such as sucrose) inside the cell and thus depends on an osmotically generated ΔΨ between cell interior and exterior. Because water stress reduces Ψxylem, a major cause for growth inhibition under water deficit may simply be the smaller ΔΨ, which reduces cell water uptake (Nonami et al. 1997). In addition, cell expansion involves cell-wall loosening, which requires acidification by protons (H+) pumped from the cell interior to the cell wall in exchange for K+ maintaining the cell’s electrical charge balance (Stiles and Van Volkenburgh 2004). It is possible that the increase in apoplast pH due to water deficit also directly interferes with cell expansion (Bacon et al. 1998). Therefore, growth rates change rapidly with fluctuating Ψxylem and pHxylem, and a reduction in shoot and leaf growth is the first visible sign of vine water deficit (Williams et al. 1994, Stevens et al. 1995). Root growth also decreases, but less so than shoot growth, which increases the root:shoot ratio of the vine and maintains a supply of water and nutrients to the shoots (Hsiao and Xu 2000). By inducing a blockage of ion channels in the stele, ABA reduces the release of K+ (and possibly other nutrient ions, such as H2PO4−, as well) from the root cortex into the xylem (Roberts and Snowman 2000, De Boer and Volkov 2003). This traps K+ taken up from the soil (ABA does not affect K+ uptake) and delivered from the shoots in the phloem and leads to K+ accumulation in the roots. The osmotic activity of K+ lowers Ψroot, which enhances phloem water import. This helps the vine to maintain root growth, while avoiding K+ transport to the leaves, where it would only worsen the water deficit by lowering Ψleaf. In addition, the roots also osmotically adjust by accumulating sugars and amino acids in order to lower Ψroot to favor water uptake and, possibly, reduce the risk of xylem cavitation (Schultz and Matthews 1988). Nevertheless, cavitation may be important in inhibiting shoot growth at moderate water deficits.
The stomatal closure (high rs) discussed above reduces water loss by transpiration but also limits photosynthesis, because CO2 diffusion into grape leaves is much more dependent on open stomata than is H2O vapor diffusion out of the leaves (Boyer et al. 1997, Flexas et al. 1998, Escalona et al. 1999). But even a stress that is mild enough not to affect photosynthesis can reduce shoot growth and hence canopy development. The combination of smaller total leaf area and decreased photosynthesis will result in reduced daily assimilate production of the vine (Perez Peña and Tarara 2004). While sugar concentrations may remain high in the leaves, starch becomes depleted and assimilate export decreases (Quick et al. 1992, Lawlor and Cornic 2002). Accumulation of solutes (mainly sugars, amino acids and, to a lesser extent, organic acids or even K) by growing organs (root tips, expanding leaves and fruits) is a common reaction to water stress. This osmotic adjustment maintains turgor and allows continued, although slower, growth (Morgan 1984). When expansion is complete (such as in fully grown leaves), tissues gradually lose this ability to osmoregulate, but leaves developing later in the season can achieve a greater capacity for osmotic adjustment than leaves formed earlier. Therefore, during water stress, older leaves wilt at higher Ψleaf than younger leaves (Patakas et al. 1997), even though young leaves may be more sun-exposed. Leaf wilting decreases the leaf surface-area and leaves hang down and become parallel to the solar rays (Smart 1974), which effectively reduces light absorption. This markedly decreases a potential energy overload that would be damaging to the photosynthetic system (Flexas et al. 1999, Lawlor and Cornic 2002). Nevertheless, water stress can be especially damaging at high light intensity with large photon fluxes.
Under severe stress, when Ψleaf decreases below approximately −1.5 MPa (depending on variety) and RWC decreases below about 75%, photosynthetic metabolism is progressively impaired (Hsiao 1973, Escalona et al. 1999). This metabolic limitation of photosynthesis is irreversible, in contrast to the stomatal limitation under mild water stress (Escalona et al. 1999, Lawlor and Cornic 2002). The depression in photosynthesis is accompanied by only a slight decrease in respiration and, as the stress becomes more severe, an increasing proportion of fixed CO2 is lost from the leaf. This CO2 is increasingly derived from stored carbohydrates (Lawlor and Cornic 2002), whose depletion is accompanied by accumulation of amino acids, particularly proline and glutamate, because their production (supply) exceeds consumption (demand). This in turn may lead to nitrate accumulation in leaves, possibly due to feedback inhibition of NR. Incidentally, it is unknown whether this mechanism may result in incorrect fertilizer recommendations based on analysis of petioles collected from water-stressed vines (in other words, to withhold N application when in fact more water should be applied). If the water stress is not relieved by rainfall or irrigation, the increased ABA and reduced cytokinin contents accelerate leaf aging and lead to senescence of older leaves (Jackson 1997, Yang et al. 2002). Senescence is accompanied by a decline in chlorophyll and remobilization of carbon, proteins, and nutrients from these leaves, and followed by leaf abscission. Abscission can also be induced by excessive xylem cavitation and enables vines to survive severe drought by drastically reducing evaporation and conserving resources. Some of the sugars, amino acids, and phloem-mobile mineral nutrients can be recycled to the fruit, which may partially sustain ripening.
Grapevines react strongly to nutrient deficit.
The viticultural literature abounds with reports of the stimulating effects of nutrient (especially N) supply on vine vigor (see Rantz 1991, Conradie 2005). However, nutrient uptake is mainly controlled by demand of the vine and thus varies according to growth requirements. Growth is the pacemaker for nutrient uptake, and limiting the supply of a nutrient ion slows down shoot growth to a rate consistent with supply (Clarkson 1985, Gastal and Lemaire 2002). That growth increases when N supply increases may merely reflect the fact that N is frequently limiting for growth. Indeed, both N uptake (Keller et al. 1995) and shoot growth (Spayd et al. 1993, Keller and Koblet 1995) of grapevines show saturation-type responses to increasing soil N. However, the distribution of available N in the soil is extremely heterogeneous. Nitrate concentrations in the soil water can vary over several orders of magnitude, both temporally and spatially, even over short distances (Robinson 1994, Crawford 1995, Keller et al. 1995, 2001b). Moreover, nutrient delivery by mass flow (Marschner 1995) and mineralization of organic N (Conradie 2005) slow down in a drying soil, so that nutrient uptake becomes increasingly difficult for grapevines (Conradie 2005, Schreiner 2005), especially if the water deficit is sufficient to slow root growth. Of course, the reduction in shoot growth induced by water deficit also decreases nutrient demand of a vine, and delivery of nitrate and K to shoots has been found to decrease in drying soil (reviewed by Davies et al. 2002, Mpelasoka et al. 2003). Nevertheless, the major effect of water deficit might be on nitrate reduction (NR activity declines rapidly and reversibly at low Ψleaf) rather than on N supply (Lawlor and Cornic 2002). It is surprising how ignorant we still are on the interactive effects of two so critically important environmental variables.
Because N is so important to them, vines have evolved mechanisms to absorb nitrate as quickly as possible. Roots react to localized sources of N (or P) by activating uptake and initiating lateral root proliferation within the nutrient-rich zones (Robinson 1994, Scheible et al. 1997, Forde 2002, Gastal and Lemaire 2002), especially when vine N status is low. Nutrient deficiency prompts a range of general stress responses in the vine, including cessation of cell division and cell expansion, changes in vine morphology, starch accumulation, decrease in photosynthesis, and modification of metabolism to adapt to limited nutrient supply (Grossman and Takahashi 2001). Specific responses to limitation of a particular nutrient include induction of transport systems to enhance uptake and remobilization of stored reserves of that nutrient. Nutrient (particularly N and P) deficiency can trigger plant responses that are very similar to effects of soil water deficit, namely increased ABA and reduced cytokinin production, increased pHxylem (increasing guard-cell sensitivity to ABA), increased rh and rs, decreased transpiration (but no effect on Ψleaf), restricted leaf expansion and senescence of older leaves, and increased root:shoot ratio (Jackson 1997, Clarkson et al. 2000, Wilkinson and Davies 2002). Although both nutrient and water stress limit growth, the inhibition may be due to different mechanisms; nutrient deficit interferes with cell division (Lemaire and Millard 1999, Gastal and Lemaire 2002), whereas water deficit primarily reduces cell expansion. Reduced influx of root-derived cytokinins in response to N deficiency probably decreases both cell division and cell expansion in shoot organs (Coruzzi and Zhou 2001, Takei et al. 2002, Forde 2002, Kakimoto 2003). Leaf expansion is particularly sensitive to fluctuations in N supply, resulting in marked changes in the leaf area:plant-weight ratio (LAR) (Keller and Koblet 1995). Root growth, on the other hand, may increase in response to N deficiency (Keller and Koblet 1995) and reduced cytokinin synthesis (Kakimoto 2003, Werner et al. 2003). Thus when nutrient supply limits growth, roots become relatively stronger sinks than shoots in order to alleviate the deficiency by improving nutrient uptake from previously untapped soil regions.
Nitrogen deficiency also limits photosynthesis (Figure 2A⇓) by reducing the activity of photosynthetic enzymes, such as rubisco (Chen and Cheng 2003). This enzymatic inhibition in turn increases rs, but also results in potential energy overload. To avoid excessive light absorption that would damage the photosynthetic apparatus, leaves decrease their chlorophyll content (Figure 2B⇓) and activate their thermal energy-dissipation and antioxidant systems (Chen and Cheng 2003). In other words, the reduced chlorophyll content of N-starved vines (Spayd et al. 1993, Keller et al. 2001b) is a consequence, not a cause, of the decrease in photosynthesis. Since inadequate N supply generally suppresses growth more than photosynthesis, carbohydrates accumulate in the leaves (Gastal and Lemaire 2002, Lawlor 2002), which often leads to anthocyanin synthesis in the petioles and leaf veins. Surplus sugar also may become available for export to the roots (Lemaire and Millard 1999) since, in contrast to the response to water deficit, it is not needed for osmotic adjustment. But similar to their reaction to water stress, leaves also decrease the angle between leaf blade and petiole, which further reduces light absorption to avoid energy overload. Another similarity to water stress is that under prolonged N stress the vine starts shedding older leaves. Abscission is accompanied by remobilization of nutrients in those leaves and redistribution to developing organs such as young leaves and fruit (Lawlor 2002). Taken together, the similarity of vine responses to water and N deficit suggests that the combined effects of these two stress factors may be mostly additive, rather than interactive. Abundant supply of water and/or N also may make vines more susceptible to stress when one of the two inputs is withheld.
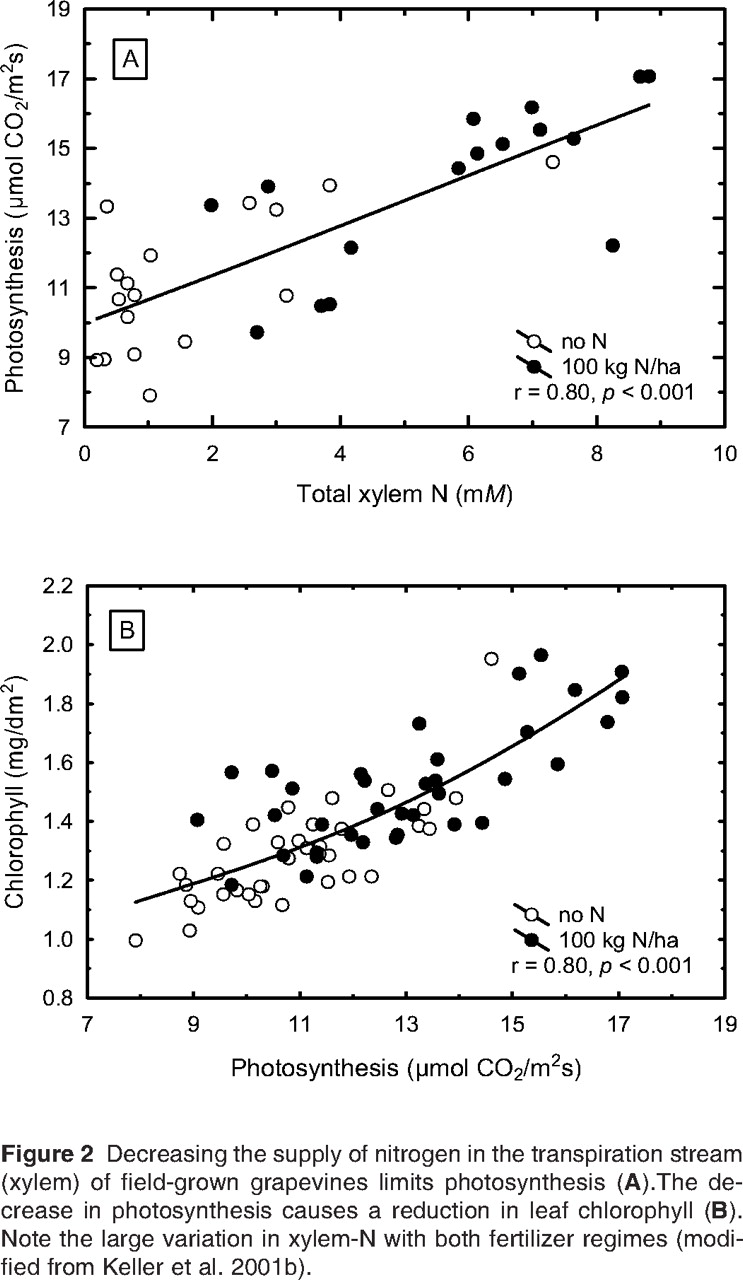
Decreasing the supply of nitrogen in the transpiration stream (xylem) of field-grown grapevines limits photosynthesis (A).The decrease in photosynthesis causes a reduction in leaf chlorophyll (B). Note the large variation in xylem-N with both fertilizer regimes (modified from Keller et al. 2001b).
Abundant N availability sometimes (Spayd et al. 1993), but not always (Keller et al. 1995, Hilbert et al. 2003), decreases P uptake. The reason could be the N-induced reduction in root carbohydrate status, which in turn might limit carbon availability for mycorrhizae. Field-grown grapevines rely heavily on mycorrhizal fungi for P uptake (Schreiner 2005), but this may not apply to short-term pot experiments. The microbial symbionts transfer P to the root in exchange for sugar, which represents the price paid by the vine for enhanced P acquisition (in effect, mycorrhizae are an extension of the root sink). Phosphate is a major substrate in energy metabolism (after all, ATP contains three phosphate groups) and synthesis of nucleic acids and membranes. It also plays an important role in photosynthesis, respiration, and regulation of a number of enzymes, thus P starvation also decreases photosynthesis. Phosphate deficiency can also increase rs and rh, so that the reduction in water supply to growing organs restricts cell expansion. This results in a strong decrease of leaf expansion, which is similar to the response to N deficit (Clarkson et al. 2000), as is the production of anthocyanins from excess sugars in the leaves. Suboptimal supply of P also limits cell division (Chiera et al. 2002), restricting leaf initiation in the shoot apical meristem and expansion of newly developed leaves and young berries. Reduced cell division may be responsible for the inhibition of cluster initiation (Skinner et al. 1988) and lateral shoot growth (Grant and Matthews 1996) in P-deficient vines. Therefore, although as a phloem-mobile nutrient P can be recycled from old leaves, P deficiency primarily restricts meristematic sink activity, and the resulting lack of demand for assimilates may be responsible for the reduction in photosynthesis and assimilate partitioning. In addition, insufficient P supply appears to restrict magnesium (Mg) transport in the xylem, which can lead to symptoms of Mg deficiency (Skinner and Matthews 1990).
Another nutrient that is important for meristem activity is boron (B). Boron differs from all other micronutrients in that it exists as a neutral molecule at physiological pH and appears to be taken up into plants by simple diffusion, especially at high external concentration (Reid 2001). Hence the roots may not be able to exclude excess B, which increases the risk of B toxicity, the symptoms of which are virtually identical to those of B deficiency. In contrast, active transport into root cells is probably involved when the concentration in the soil water is low (Reid 2001). Insufficient B availability rapidly results in cessation of cell division, whereas cell expansion appears to be unaffected by B deficiency (Clarkson and Hanson 1980). Nevertheless, the inhibition of cell division leads to stunted shoot and root growth (Gärtel 1993). Symptoms are usually confined to young tissues, since B is phloem-immobile and thus cannot be recycled from old leaves. Bushy, branched shoot growth and poor bud fruitfulness may be a carryover effect of insufficient B available for proper primordium formation in the previous season (Gärtel 1993). In addition, reduced N uptake in B-deficient vines leads to low leaf-N status and sugar and starch accumulation in the leaves (Camacho-Cristóbal and González-Fontes 1999). The importance of B for pollen tube growth leads to poor fertilization as a consequence of B deficiency. This results in poor fruit set and a condition termed “hen and chicken” (French millerandage) with clusters containing small, seedless berries (Gärtel 1993).
Growth versus reproduction: Do we have a winner?
It is commonly held that vegetative growth of grapevines is more sensitive to water stress than is reproductive growth (Williams et al. 1994), but this may not be true for all developmental stages of the vine. Similarly, the extent to which reproductive growth is affected by limited N availability varies with developmental status. Both water and nutrient deficits typically reduce yield, particularly if the deficit occurs early in the season (Williams and Matthews 1990). Nevertheless, shortly after budbreak the developing flower clusters compete successfully with the growing shoots for limited water. Once inflorescences contribute assimilates via their own gas exchange (Leyhe and Blanke 1989), they may survive even relatively severe drought sufficient to stunt shoot growth (M. Keller, unpublished data). Closer to and during bloom, however, the metabolic requirements of flowers far exceed their own contribution (Blanke and Leyhe 1989). Even relatively mild water deficit can interfere with pollen production, pollination, and fertilization, which results in poor fruit set or abscission of inflorescences (Smart and Coombe 1983, Callis 1995). Providing water to only a portion of the roots of pot-grown Syrah during bloom reduced fruit set, although no symptoms of water stress were visible on those vines (Rogiers et al. 2004). The effect of bloom-time water deficit is exacerbated by N deficiency and could also reduce yield potential (bud fruitfulness) for the following season, if the stress results in poor cluster initiation. Water stress normally reduces cluster initiation, but this is not always the case (Smart and Coombe 1983, Williams and Matthews 1990, Williams et al. 1994). A mild water deficit decreasing canopy density may actually increase initiation due to the improved canopy microclimate. Mineral nutrition also influences cluster initiation, but little is known about the cause-effect relationships. They likely involve changes in microclimate in addition to direct effects of nutrients on primordia formation. Fruit set of abundantly irrigated pot-grown Müller-Thurgau was unaffected by N supply (Keller and Koblet 1994), whereas maximal cluster initiation required an optimum (rather than maximum) supply of N (Keller and Koblet 1995). Water deficit during bloom of pot-grown Cabernet Sauvignon reduced fruit set only in those vines that were also N deficient but not in vines that received supplemental N (Keller et al. 1998). Similarly, prebloom application of N fertilizer to deficit-irrigated Riesling vines (Spayd et al. 1993) and to Müller-Thurgau vines growing in a vineyard with a complete floor cover (Keller et al. 2001a, Figure 3⇓) improved fruit set compared with vines that did not receive fertilizer. The effects of water and N deficit are likely mediated by a combination of low Ψinflorescence and changes in xylem-sap pH, ABA, and cytokinins in addition to assimilate supply via the phloem. Moreover, cavitation in the inflorescence xylem may not be as easy to repair as cavitation in leaves, but none of these possibilities has been investigated.
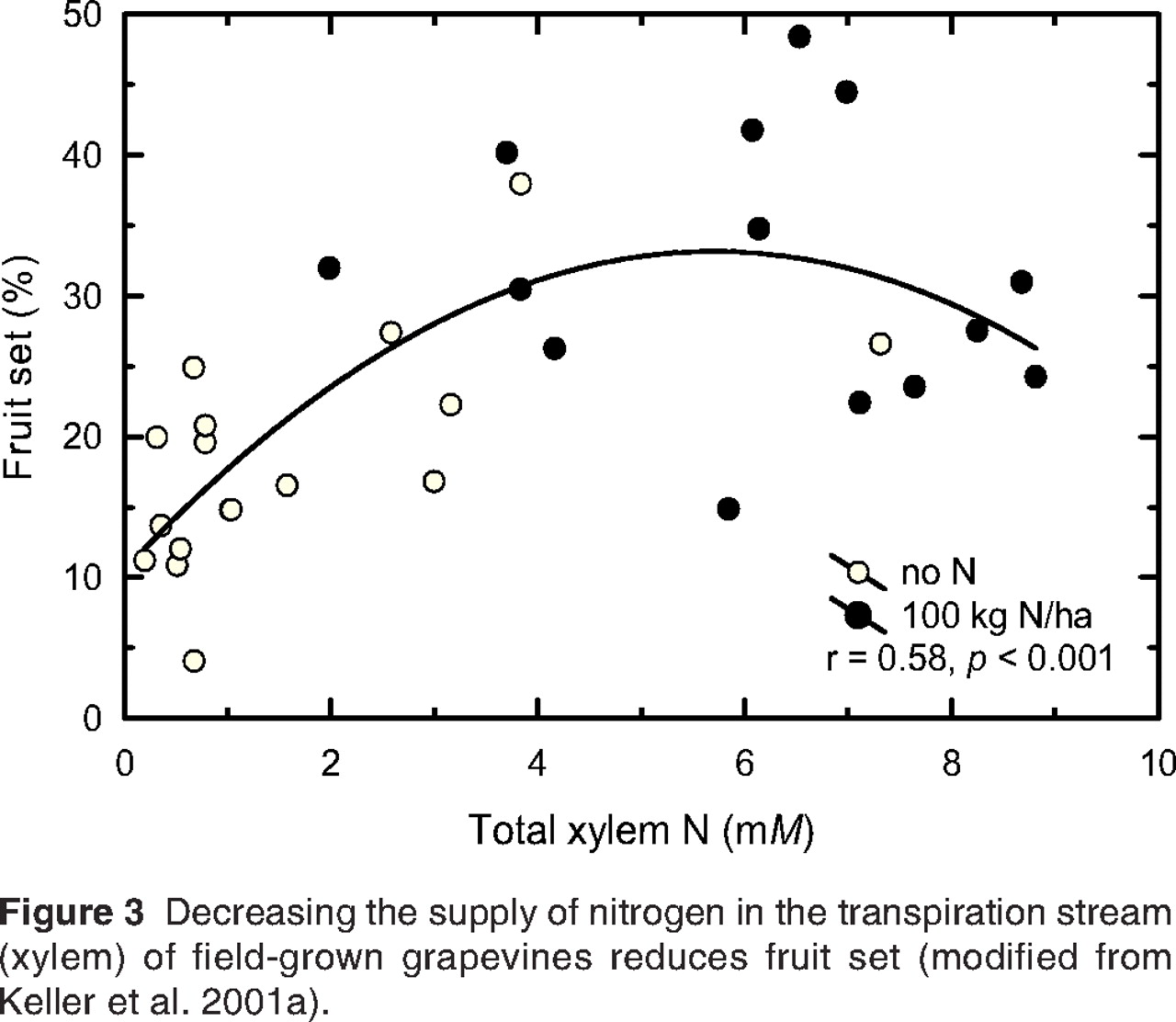
Decreasing the supply of nitrogen in the transpiration stream (xylem) of field-grown grapevines reduces fruit set (modified from Keller et al. 2001a).
When a stress occurs after fruit set, grapevines generally maintain fruit growth and ripening at the expense of shoot and root growth (Williams et al. 1994) and replenishment of storage reserves in the roots (Smith 2004). This can, of course, have long-term implications, because the vine is heavily dependent on stored reserves for budbreak in the following season. A small root system supplying water to a large canopy could moreover result in a very steep ΔΨsoil/leaf, which could render vines vulnerable to xylem cavitation. A decrease in root:shoot ratio due to fertilizer addition (Keller et al. 1995, Zerihun and Treeby 2002) can further increase a vine’s susceptibility to drought, particularly in irrigated vineyards in the event that irrigation is withheld (Sperry et al. 2002). In nonirrigated plants, however, fertilizer application may increase the resistance to xylem cavitation, which tends to counter the lower root:shoot ratio, although excessive N supply can reduce water uptake due to the buildup of solutes in the soil water (Keller and Koblet 1994, Sperry et al. 2002).
The final weight of grape berries is determined by the number of cell divisions before and after bloom, the extent of expansion of these cells, and the degree of shrivel prior to harvest. Limited water supply during the period of cell division and expansion restricts berry enlargement (limiting berry size). In keeping with the effect on shoot growth, reduced Ψxylem and pHxylem could also be partly responsible for the smaller berry size of water-stressed vines, particularly if the stress occurs before veraison. The same extent of water deficit occurring during the postbloom cell division phase of berry growth normally reduces berry size more than if it occurs after that phase (Hardie and Considine 1976, Williams and Matthews 1990, McCarthy 1997). Applying more water later in the season cannot compensate for the decrease in berry size due to early-season deficit, possibly because limited xylem flow into the berry after veraison (Düring et al. 1987, Findlay et al. 1987, Rogiers et al. 2001, Ollat et al. 2002) strongly decreases the sensitivity of berry water status to soil and plant water status (Greenspan et al. 1994, 1996). The response of berry growth to water deficit also depends on the crop load (fruit mass per unit vine leaf area). Berry growth may be reduced more on vines bearing a heavy crop than on vines with a light crop load, because reduced sugar availability (per berry) and import into the berries also decreases the ΔΨ driving water uptake by those berries (Fishman and Génard 1998). In contrast to water deficit, when N supply is limited after bloom, berries may actually grow larger (Keller et al. 1998, Hilbert et al. 2003, Rogiers et al. 2004). This is somewhat surprising but may be due to limited competition from growing shoot tips. More sugar available for import into the berries would also increase berry water uptake so that berry size increases. Even though this N effect may not always carry through to harvest, it might be possible to further limit berry size by applying N to vines that are experiencing postbloom water deficit. However, rates of N fertilizer required for an optimum response have yet to be determined, and there is always a danger of reducing fruit quality by N oversupply. Of course, if stress is imposed early enough to reduce fruit set (and hence crop load), the size of the remaining berries may increase and partially compensate for the loss in yield potential, which would offset potential benefits of smaller berry size.
Fruit Composition and Implications for Irrigation and Nutrition Strategies
Large, dense canopies that result from abundant water and nutrient availability are associated with reduced fruit sugar, high acidity, and poor color (Jackson and Lombard 1993, Dry and Loveys 1998). It is generally accepted that some form of water and nutrient deficit is beneficial for fruit composition and wine quality, as long as the stress is not too severe. After all, grapes have been grown successfully on relatively marginal sites for centuries, and controlled application of stress is the premise of modern deficit irrigation strategies and reduced fertilizer application. While significant gains in terms of fruit composition may be due to indirect effects of smaller, more open canopies and, sometimes, reduced crop load, there may also be more direct effects of water and nutrient deficits on berry size and composition. Mild to moderate stress generally seems to be most effective if applied during the first phase of rapid berry expansion by limiting growth and reducing canopy density (Dry et al. 2001). Solute transport in the phloem can continue at low Ψleaf, so that berries grown on mildly water-stressed vines are not only smaller, but also often have higher sugar concentration. Sugar accumulation has been reported to be less sensitive to water stress than berry growth (Matthews and Anderson 1988, Williams and Matthews 1990, Stevens et al. 1995), although this could simply be due to a decrease in the ability of berries growing under limited sugar supply to accumulate water, so that the sugar concentration but not the amount per berry remains constant. Moreover, the decrease in photosynthesis and sugar export from the leaves under more severe stress can lead to a reduction in berry sugar accumulation (Quick et al. 1992, Rogiers et al. 2004, Smith 2004). Postveraison water deficit may also slightly reduce berry size and improve fruit composition (Kennedy et al. 2002, Ojeda et al. 2002), although decreased fruit quality has also been reported (Dry et al. 2001), especially with severe water stress (Hardie and Considine 1976). Despite poor fruit set due to N deficiency, fruit sugar may be inadequate, because the smaller leaf area and inefficient photosynthesis limit sugar production (Keller et al. 1998).
Soil moisture and N status have little effect on berry tartrate, but malate tends to decline with decreases in both soil moisture (Stevens et al. 1995) and N availability, although the latter response may be modified by other environmental variables (Spayd et al. 1994, Keller et al. 1998). The reduction in malate is more pronounced when the deficit occurs before veraison than after veraison (Williams and Matthews 1990). The improved color of red grapes often observed with mild water stress is, in part, simply due to smaller berry size increasing the skin:pulp ratio, and to better fruit exposure. But there also may be a direct effect of water deficit enhancing the production of anthocyanins (Dry et al. 2001), although this has not been observed when the deficit was imposed after veraison (Kennedy et al. 2002). While accumulation of flavonols (quercetinglycosides and relatives) and hydroxycinnamates (tartrates of caffeic, coumaric, and ferulic acids) seems to be little affected by water deficit (Keller and Torres-Martinez 2004), the stimulation of the production of secondary metabolites, such as phenolics, by N deficiency is well known. Accordingly, pigmentation of Cabernet Sauvignon and Merlot grapes was maximized with low to moderate N availability (Keller et al. 1998, Hilbert et al. 2003), and maximum coloration in Syrah resulted when limited N supply was combined with water deficit between fruit set and veraison (Wade et al. 2004). It is not only total color that is affected by N, but also the distribution of individual pigments. Conditions favoring color accumulation may also lead to the most balanced anthocyanin profile (Keller et al. 1998).
Water and N availability also influence the amount and composition of nitrogenous compounds in grapes. Import of N compounds into the berry can occur in both the xylem (predominantly in the form of glutamine and nitrate) and the phloem (mostly glutamine) before veraison, but is largely restricted to the phloem after veraison. Nevertheless, substantial amounts of N can move into the berry after veraison from both newly root-absorbed N and N recycled from the vegetative parts (Rodriguez-Lovelle and Gaudillère 2002, Schreiner 2005). Therefore, berries remain responsive to N supply during ripening, which contrasts with the influence of water supply. Once inside the berry, glutamine can be converted to other amino acids, such as arginine and proline, which are the two major amino acids in ripe grape berries (van Heeswijck et al. 2001). The yeast-assimilable arginine can be accumulated throughout berry development, although in some varieties accumulation appears to cease at veraison. In contrast, most of the proline (which yeast cells cannot metabolize) is accumulated after veraison, perhaps to protect berry cells from osmotic stress caused by the accumulating hexose sugars (van Heeswijck et al. 2001). Synthesis of arginine is very limited under both water and N deficit, but reacts strongly to increasing supply, whereas proline is less affected by water and N availability (Löhnertz et al. 2000, Wade et al. 2004). Thus yeast-assimilable N increases as N availability increases (Treeby et al. 2000, Rodriguez-Lovelle and Gaudillère 2002).
The findings that developing grape berries become increasingly insensitive to water deficit (McCarthy 1997) but may remain responsive to N supply (Rodriguez-Lovelle and Gaudillère 2002) have implications for irrigation and nutrient management. Stress should be avoided before fruit set, whereas the time between fruit set and veraison is the period when shoot growth and berry size can be most effectively controlled by water deficit. This principle is exploited by regulated deficit irrigation (RDI), where a short duration of water deficit is applied as soon as possible after fruit set. During the RDI period, the soil profile is allowed to dry down until control of shoot growth has been achieved (Dry et al. 2001, Kriedemann and Goodwin 2003). Obviously, this is possible only in areas with sufficiently low seasonal rainfall or high evaporative demand and on soils with limited water storage capacity. Once shoot growth stops and especially after veraison, vines are stressed only sufficiently to discourage new shoot growth. At the end of the season the rootzone is refilled to field capacity.
When roots of a vine experience both wet and dry soil, the reaction of shoot growth may depend on whether the difference in soil moisture is perceived by the same or different roots. For instance, Eucalyptus shoot growth was not reduced when the surface soil dried out, as long as the roots had access to subsoil water (Phillips and Riha 1994). However, when separate roots of the same grapevine experienced dry and wet soil columns, shoot growth was suppressed (Dry et al. 2001, Lovisolo et al. 2002). This principle is used in another deficit irrigation method termed partial rootzone drying (PRD), in which water is supplied alternately to only one side of a vine, while the other side is allowed to dry down. This strategy attempts to separate the physiological responses to water stress (for example, ABA production, increased rs) from the physical effects of water stress (low Ψleaf, for example) (Dry et al. 2001, Davies et al. 2002). The ABA produced by the drying roots induces partial stomatal closure and reduced shoot growth, while the fully hydrated roots maintain a favorable plant water status (no decrease in Ψleaf). The wet roots also sustain the drying roots by supplying water to them (Stoll et al. 2000). In addition to increased ABA production, the drying roots produce lower amounts of cytokinins, which inhibits lateral shoot growth (Stoll et al. 2000). Most of the reduction in canopy size of vines subjected to PRD may be due to inhibition of lateral shoot growth (Dry et al. 2001). Provided the vine’s water status is maintained with PRD, berry size and yield are also maintained, while the lower canopy density often results in improved fruit quality (Dry et al. 2001, Wade et al. 2004). This is in contrast to RDI, which typically reduces berry size and yield. Alternating the dry/wet zones prevents adaptation by the vine and ensures continued production of ABA by the drying part of the root system. However, the PRD effect may be lost if the switching interval is too long, the wet soil volume too small for a given vine size, two discrete root systems cannot be established (such as following the conversion of furrow- or sprinkler-irrigated vineyards), or in poorly structured clay soils (Smart 2004).
The fundamental difference between RDI and PRD is that with RDI soil-water deficit is applied over time, whereas with PRD the deficit is applied over space. Provided seasonal rainfall is low enough, RDI always results in a plant-water deficit, while PRD usually does not if managed properly (Kriedemann and Goodwin 2003). Although PRD can be targeted to a particular growth stage, it is normally maintained over the entire season. In Australia, where PRD was developed, this method has resulted in water savings of up to 50% compared with conventional, nondeficit irrigation. Drip irrigation is generally the technique of choice, but under-vine (micro-)sprinklers, furrow, and flood irrigation have all been used successfully with both RDI and PRD (Dry et al. 2001, Kriedemann and Goodwin 2003). An additional response of vines to PRD appears to be a shift of root growth to deeper soil layers (Dry et al. 2001), whereas under standard drip irrigation, roots are often concentrated in the surface soil, even after years of exposure to RDI (J. Perez Peña, unpublished data). Evidently this adaptive behavior makes vines more drought-resistant, but it also interferes with the application of the PRD principle on soils with a deep root zone and ready access of the roots to subsurface water. Various combinations of RDI and PRD can also occur naturally in vineyards and contribute to within-vineyard variability, although in nonirrigated vineyards the severity of water stress tends to increase as the season progresses.
Since deficit irrigation strategies differ in their effect on plant water status, they should also be expected to trigger different growth responses. Decreased cytokinin production by drying roots should limit cell division in growing meristems with both RDI and PRD, but only RDI should reduce cell expansion via its effect on Ψ. Deficit irrigation and limited N supply can restrict shoot growth at any time of the season, because both production of new cells and expansion of those cells are required for continued shoot growth. Conversely, while RDI may also limit berry size throughout most of berry development, PRD should do so predominantly during the cell division phase, and low N status could even increase berry size by strongly limiting shoot growth. This might be interesting for managing irrigation in white grapes, since berry size (skin:pulp ratio) is far less important in white grapes than in red grapes, simply because skin components are not usually extracted during white winemaking. However, the high flavonol content of sun-exposed grapes (Price et al. 1995, Spayd et al. 2002, Keller and Torres-Martinez 2004) may lead to noticeable astringency or even bitterness in wine (Gawel 1998). Therefore, RDI may be less applicable for white varieties, whereas PRD may be more suitable, as long as the reduced canopy size does not lead to overexposed fruit.
Although local increases in root nitrate uptake can compensate for local soil-N deficit (Scheible et al. 1997), a reduction in shoot growth similar to that in response to PRD is often observed when N or P is supplied only to a portion of the roots (Baker and Milburn 1965, Robinson 1994). This is an area that deserves closer attention in grapevines, since combining RDI or PRD with appropriate fertigation strategies could provide a powerful tool to control shoot growth while maintaining water and nutrient supply to the fruit. The effects of water and N availability seem to be mostly additive (Rogiers et al. 2004, Wade et al. 2004), so deficit-irrigated vines may require a somewhat higher N supply to achieve photosynthetic rates as high as those of fully irrigated vines (Alleweldt et al. 1984). Vines with an optimum supply of water and N also make longer use of the growing season by extending the effective life of leaves (Lawlor 2002). However, broadcasting N fertilizer in deficit-irrigated vineyards may result in N deposition in areas inaccessible to vine roots due to the vines’ restricted rootzone. When RDI was combined with low N supply via fertigation, Syrah berry color was maximized and Botrytis bunch rot minimized, but arginine concentration was drastically reduced (Wade et al. 2004), making the resulting musts prone to sluggish or stuck fermentation (Spayd et al. 1995). In contrast, PRD combined with low N addition resulted in equally good color and low rot without the negative impact on arginine (Wade et al. 2004). Perhaps we should be speaking of regulated deficit nutrition (RDN) in analogy to the irrigation technique. Applying less N than the amount required for maximum growth early in the season can be used to control shoot growth and manipulate berry composition, while N addition later in the season might be useful to increase juice arginine where necessary. However, minor negative effects of N addition at veraison on fruit composition and bunch rot have also been reported (Christensen et al. 1994).
Cover crops or green floor covers affect grapevines in a manner comparable to deficit irrigation, because the covers use water and reduce nutrient availability for the vines (Schaller 1991, Löhnertz et al. 2000, Morlat and Jacquet 2003). Performance of vines with a permanent floor cover in a cool-humid climate was remarkably similar to that normally observed with RDI in warm-dry climates; notably reduced vigor, smaller berry size, improved fruit composition, and reduced bunch rot (Keller et al. 2001a). Vineyard floor vegetation is desirable for many reasons (Bugg and Van Horn 1998) but needs to be managed carefully to minimize competition during critical periods. A strategy termed “adaptive nitrogen management” has been developed to optimize N nutrition while minimizing fertilizer inputs into the vineyard (Perret et al. 1993). This strategy aims to manage floor covers in a way that synchronizes N availability in the soil (supply) with uptake by the vine (demand). Under cool-humid climatic conditions nutrients can be fixed in the green cover during periods of low demand and released by mowing or tilling during periods of high demand. This is useful in situations with frequent summer rainfall, but could cause problems in more arid environments. However, regions that rely on irrigation because of low summer rainfall may have better control over nutrient supply, because careful irrigation management may prevent leaching, especially of nitrate, that afflicts regions with abundant seasonal rainfall (Schaller 1991). Fertilization may be inefficient when intense rainfall or abundant irrigation leach nutrients out of the rootzone before they can be taken up by the vines. Of course, since withholding water also interferes with mineral nutrient supply, fertilizer applications may also be inefficient when lack of soil moisture impedes nutrient uptake (Löhnertz et al. 2000, Conradie 2005, Schreiner 2005). Small-scale and short-term variations in soil moisture and nutrient availability may be equally as important for vine performance and fruit composition as variations in aboveground factors, such as light and temperature. Vineyard management strategies will need to address these below-ground components of the production system in more detail to boost viticulture to the next level of quality and sustainability in the face of climate change, declining resource availability, and increasing consumer pressure. Future research should aim at integrating irrigation, nutrition, and floor covers in site-adapted soil management strategies that are tailored to the intended use of the grapes produced on each site.
Footnotes
This paper was first presented at the ASEV Annual Meeting, 29–30 June 2004, San Diego, California and published in the Proceedings of the Soil Environment and Vine Mineral Nutrition Symposium. L.P. Christensen and D.R. Smart (Eds.), pp. 91-107 (2005).
From the ASEV Soil Environment and Vine Mineral Nutrition Symposium
- Copyright © 2005 by the American Society for Enology and Viticulture
Literature Cited
Vol 56 Issue 3








{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
More from this TOC section
Similar Articles
