


Abstract
A key objective of this review is to describe the tissue types in the grape berry from a phenolics point of view, that is, an account of cell types based on the phenolic components they contain. To complement this, the major soluble phenolics of the grape are described with an emphasis on the relative abundance of some of the components in each class. The intent of this approach is that the juxtaposition of tissue types and phenolic composition will help identify features of both that influence extraction during winemaking.
Berry Anatomy and Phenolic Composition
The skin of a grape berry contains tannins and pigments, the pulp contains juice but no pigments, and the seeds contain tannins, as shown in Figure 1⇓. While this figure is a serviceable representation of the grape, it lacks sufficient resolution to provide any insight into how phenolics are extracted from fruit during winemaking.

A simple diagram of the grape berry used to describe the tissues and phenolic composition in an introductory wine class.
Considine and Knox (1979) published an excellent account of grape berry pericarp development and distinguished two cell types in the skin: the single layer of clear epidermal cells on the outside of the fruit and six hypodermal layers beneath the epidermis. The exact number of hypodermal layers depends on the variety (Hardie et al. 1996). The pulp was described as having three easily distinguished tissues: the outer mesocarp, the inner mesocarp, and the vascular tissue separating the inner and outer mesocarp. The vascular tissue in turn is comprised of xylem, phloem, and vascular parenchyma.
The structure denoted as the seed coat (testa) has two very distinct layers, both of which are derived from the outer integument of the ovule during seed development. From a phenolics standpoint, these two layers could not be more different. The inner cell layers of the outer integument undergo a series of periclinal divisions (parallel to the surface of the seed) that results in a layer of cells that are thin in the fossetts of the seed but much thicker in the beak. Near veraison, when the seed coat still appears yellow, the innermost cells of the outer integument undergo lignification. The cells outside the lignified layers retain their thin primary cell walls except for the outermost ones. The outer walls of these cells thicken to form a cuticle on the surface of the seed. The thin-walled cells between the cuticle and the inner lignified layers are perhaps the most important in terms of phenolics because they contain nearly all of the soluble seed phenolics. The familiar seed browning that accompanies fruit ripening after veraison is thought to be the result of tannins and flavan-3-ols in this layer undergoing oxidation. Thus the browning of seeds during ripening is not associated with the process of lignification, but instead is attributed to oxidation of the polyphenols in the thin-walled cells that lie outside the lignified cells of the hard seed coat. During initial growth of the seed, the polyphenols accumulate along with fresh weight and both reach a maximum around veraison. The seed undergoes water loss even though it is encased in a fleshy pericarp, thus “drying” in a fleshy fruit. Seed drying and the accompanying browning occurs after veraison, during the period of ripening when sugars accumulate and malic acid is metabolized in the skin and mesocarp. It has been shown that the appearance and color of the seed coat might be a useful indicator of berry ripeness in Shiraz (Ristic and Iland 2005).
The study of pericarp development after anthesis reported by Hardie et al. (1996) is particularly valuable in understanding the nature of cells making up the hypodermis (skin) and mesocarp (flesh). Fourteen days after anthesis the hypodermal cells of Traminer had a mean radial cell width of 7.0 ± 0.5 m and a cell wall thickness of 0.2 ± <0.1 mm. At this time the cells of the inner mesocarp were already larger than the hypodermal cells, having a mean radial cell width of 17.8 ± 1.5 mm. However, the cell wall thickness of the inner mesocarp cells was 0.17 ± <0.1 mm, similar to that of the hypodermal cells 14 days postanthesis. After the cell divisions and cell expansion associated with berry development (126 days postanthesis), the radial cell width of the hypodermal cells declined from 7.0 ± 0.5 mm to 5.4 ± 0.6 mm while the mesocarp cells expanded from 17.8 ± 1.5 mm to 68.0 ± 4.5 mm. Thus, by harvest the mesocarp cells had a radial width 9.7 times greater than the hypodermal cells of the skin. As the width of the hypodermal cells declined and as the mesocarp cells expanded, important changes occurred in the cell walls of these two tissues. From 14 to 126 days postanthesis, the thickness of the hypodermal cell walls increased nearly 10-fold from 0.2 ± <0.1 mm to 1.9 ± 0.2 mm. During this same time the cell walls of the mesocarp became 59% thinner, from 0.17 ± <0.1 mm to 0.1 ± <0.1 mm. Thus, in fruit at harvest hypodermal cells of the skin had cell walls nearly 20 times thicker than the cell walls of the mesocarp cells. An estimate of relative cell volume based on values provided by Hardie et al. (1996) for cell dimensions shows that the thin-walled cells of the mesocarp are about 75 times larger than the very thick-walled hypodermal cells. This representation of berry pericarp at harvest is important for two reasons. First, these two tissues have remarkably different phenolic composition at harvest, and second, the difference in cell size, and in particular the difference in cell wall thickness, is crucial for understanding the more practical question of how phenolic compounds in these tissues are extracted during winemaking.
Most work on the polyphenolics of grape berries has focused on those that are soluble in aqueous or organic solvents. From a biological perspective, the insoluble cu-tin of the epidermis and the insoluble lignin of the hard seed coat are just as phenolic and just as important as the tannins and pigments in the skin and the tannins in the seed. Nevertheless, because of their importance in winemaking, this overview will consider only the soluble phenolics of the grape berry and will address them in the order of their abundance.
Phenolic Classes in the Grape Berry
Tannins.
The most abundant class of soluble polyphenolics in grape berries are the polymeric flavan-3-ols (tannins) found in the hypodermal layers of the skin and the soft parenchyma of the seed between the cuticle and the hard seed coat. A hypothetical tetramer made of the subunits that comprise most grape tannins is shown in Figure 2⇓, where catechin, epicatechin, and epigallocatechin are “extension” subunits because they share an interflavan bond between their carbon 4 position and carbon 8 of the adjacent subunit. Epicatechin gallate is the terminal unit and has its carbon 4 position free but has gallic acid esterified to the hydroxyl group on carbon 3. Catechin and epicatechin make up most of the extension units in grape tannins, with epicatechin usually being the more abundant of the two.

This hypothetical condensed tannin is made up of four subunits: catechin, epicatechin, epigallocatechin, epicatechin gallate. The first subunit (catechin) is bound to the second (epicatechin) by an interflavan bond between carbon 4 of catechin and carbon 8 of epicatechin. The first three subunits are referred to as “extension” subunits and only the epicatechin gallate has a free 4 position and is thus referred to as a “terminal” unit.
The tannins of grape are actually a very diverse set of biomolecules. They vary in size from dimers and trimers up to oligomers with more than 30 subunits. Skin tannins differ from seed tannins in several important ways. First, the average size of skin tannins (expressed as their mean degree of polymerization: mDP) is much larger than seed tannins, and skin tannins contain epigallocatechin subunits whereas seed tannins generally lack epigallocatechin. However, the smaller seed tannins usually have a higher proportion of their subunits as epicatechin gallate whereas epicatechin gallate is usually not present in skin tannin (Cheynier 2005). This difference in signature subunits has been used to estimate the relative proportions of skin tannins and seed tannins in Pinot noir wines (Peyrot des Gachons and Kennedy 2003).
Anthocyanins.
The anthocyanins are co-located with tannins in the thick-walled hypodermal cells of the skin. Anthocyanidins found in grape are shown in Figure 3⇓. A variety such as Pinot noir that lacks acylated pigments has only the basic five anthocyanins whereas varieties that produce all three types of acylated derivatives could have as many as 20. This diversity is multiplied further in vivo because at vacuolar pH there are two distinct chemical forms of each anthocyanin, the neutral quinonoidal anhydro base and the flavylium cation form (Figure 3⇓). The ratio of these two chemical forms of anthocyanin is highly pH dependent with a pKa of ~4.

The anthocyanins found in grape berries.
Hydroxycinnamates.
The third most abundant class of soluble phenolics in grape berries are the hydroxycinnamates, which are present in hypodermal cells along with tannins and anthocyanins and in mesocarp and placental cells of the pulp. Thus, while they are found in red wines, they are usually the most abundant class of phenolics in free-run juice and consequently in white wines. The predominant hydroxycinnamates present in grape berries are shown in Figure 4⇓. They are present almost entirely as the trans isomers but small amounts of the cis isomers have also been reported.

The hydroxycinnamates found in grape berries.
Flavan-3-ols.
The flavan-3-ols are represented by catechin and epicatechin (Figure 5⇓). They differ around the two stereo centers of the flavan-3-ols, catechin having the 2,3-trans configuration and epicatechin being 2,3,-cis. These features of the molecules become important in considering their biosynthesis below.
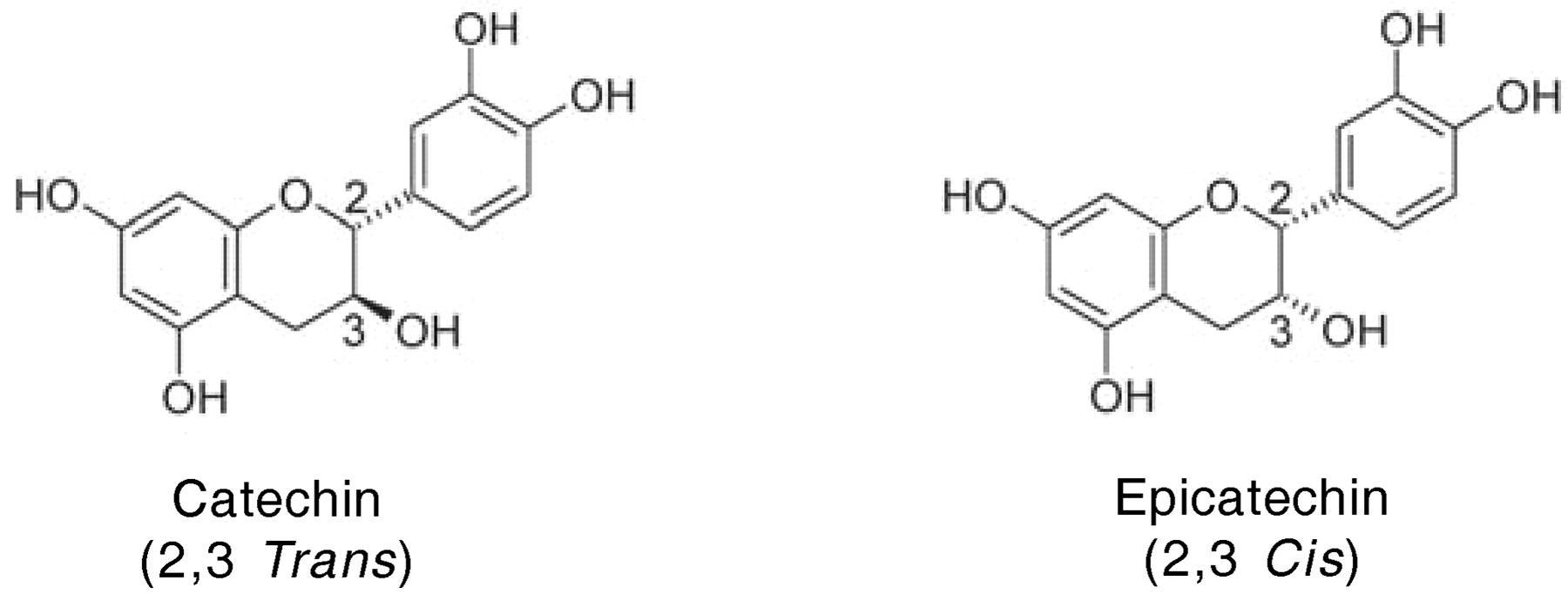
Structure of catechin and epicatechin, the most abundant monomeric flavan-3-ols in grape.
The chemical structure of the flavan-3-ols, catechin and epicatechin, compared to the known structures of tannin polymers suggests a precursor product relationship; that is, the monomeric flavan-3-ols are widely considered to combine to give tannins. However, the way in which the 4–6 and 4–8 interflavan bonds are formed remains an important unknown in plant biology. Indeed there are at least two closely related problems in our understanding of tannin biosynthesis from the corresponding flavan-3-ols. The first is how interflavan bonds are formed and in what compartment of the cell the reaction takes place. If it occurs in the vacuoles where the tannins are finally located, then the issue of how they are transported across the tonoplast does not arise. However, if the interflavan bonds are formed in the cytoplasm or in organelles such as plastids, then the question of how tannins are transported across membranes becomes an important one. It has recently been shown that mutants of Arabidopsis with a defective plasma membrane H+-ATPase are unable to accumulate tannin in vacuoles of the seed coat endothelial cells (Baxter et al. 2005), suggesting that tannin accumulation requires proper endomembrane trafficking and that at least one of the requisite H+-ATPases plays a specific role in tannin biogenesis and vacuole formation. This in turn suggests that understanding how tannins are transported across membranes is essential to knowing how they accumulate in grape skin hypodermal cells and the soft parenchyma of the outer seed coat.
Flavonols.
The flavonols kaempferol, quercetin, myricetin, and isorhamnetin are found in wines, but in the berry they are present as the corresponding glucosides, galactosides, and glucuronides (Figure 6⇓). In Pinot noir, Shiraz, and Merlot fruit, the amount of these compounds has been shown to be highly dependent on light exposure of the tissues in which they accumulate (Price et al. 1995, Spayd et al. 2002, Downey et al. 2004). In Pinot noir, berries on the sun side of a cluster had higher levels than berries from the shade side. This heterogeneity extended to individual berries where skin from the sun side of an exposed berry had higher levels than the shade side of the berry (Price et al. 1995). The variation in flavonol levels among berries on a cluster and clusters on a vine probably accounts for the wide range of flavonols reported in wines and fruit. In most cases they are less abundant than all of the phenolic classes considered above.

The flavonols of grape: found in the berry as glucosides, galactosides, and glucuronides where R3 is glucose, galactose, or glucuronic acid, respectively.
Stilbenes.
A complete account of all grape phenolics would have to include the stilbenes and the closely related viniferins, which are produced by the fruit and the vegetative tissue in response to fungal attack and exposure to high levels of UV light (Jeandet et al. 1991). The amount of stilbenes in fruit at harvest is thus understandably variable and the quantities are usually even far less than the flavonols. Despite the great interest in these compounds for their supposed effects on human health, they will not be considered further in what follows because of their low abundance.
Phenolic Biosynthesis in Grape Berry Tissue
Descriptions of phenylpropanoid biosynthesis usually begin with the amino acid phenylalanine, which is a product of the shikimate pathway. The shikimate pathway begins with erythrose-4-phosphate and phosphoenol pyruvate and is responsible for producing the other aromatic amino acids tyrosine and tryptophan. It is thought that nearly all plant cells must be able to produce these aromatic amino acids because they are required for protein synthesis. The amount of carbon that must be allocated to the shikimate pathway in grape berry cells that do not accumulate phenylpropanoids is minimal because the amount of protein in berry cells is very low. However, in cells that become lignified, such as the xylem of the vasculature and the hard seed coat, a large amount of carbon must be allocated to the shikimate pathway to provide phenylalanine. Likewise, cells that accumulate the soluble phenolics discussed above must also allocate carbon to the shikimate pathway in proportion to the total amount of polyphenolics they produce and accumulate.
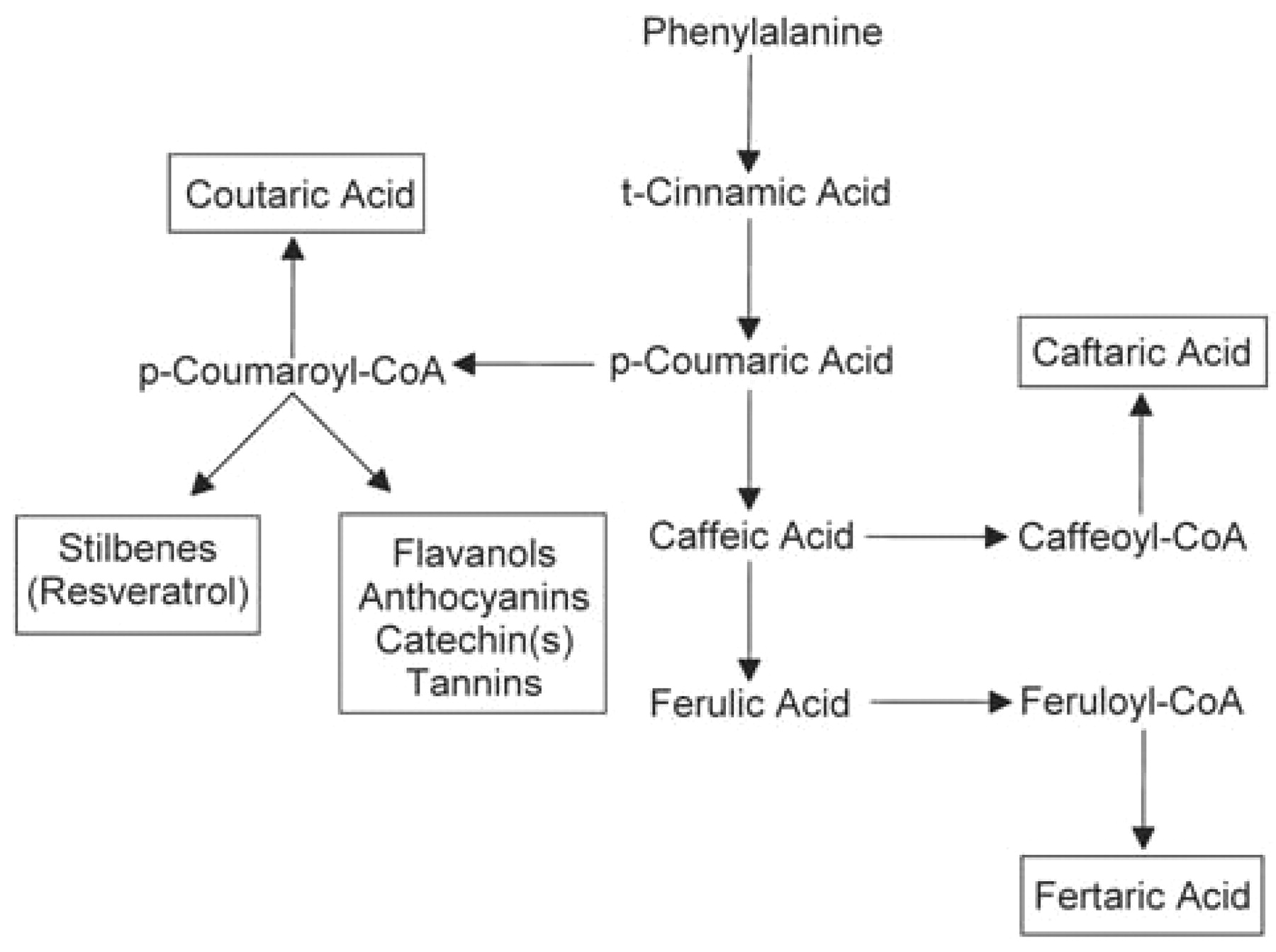
An outline of phenolic biosynthesis beginning with phenylalanine is shown in Figure 7⇓, with compounds that accumulate as soluble products in grape berry cells outlined. The amount of phenylalanine, free cinnamic acids, and their CoA esters are reported to be extremely low in tissues where care is taken to prevent ester hydrolysis prior to analysis. Only anthocyanins in the skin appear to behave anything like typical end products and seem to show little turnover once formed. The other phenolic classes exhibit patterns of accumulation and subsequent decline during ripening, suggesting their degradation, utilization in biosynthesis of other compounds, or covalent association with other cellular components (which prevents normal solvent extraction). In all cases much more is known about how various classes of grape polyphenolics are produced than about what becomes of them when they show a decline per berry during ripening.

The biochemical pathway that leads to the major soluble phenolic classes found in the grape berry. Compounds or classes that accumulate in the fruit are outlined by rectangles and intermediates (not outlined) are usually present at very low levels. Caftaric, coutaric, and fertaric acids are the tartrate esters of the respective cinnamic acid.
The origin of only the soluble phenolics in grape berries is shown in Figure 7⇑, but in cells producing lignin the free cinnamic acids and their CoA esters are converted to p-coumaryl alcohol, coniferyl alcohol, and sinapyl alcohol by a family of oxygen dependent cytochrome P450 hydroxylases and S-adenosylmethionine-dependent methyl-transferases. These alcohols are the substrates for the polymerization reactions that produce lignin.
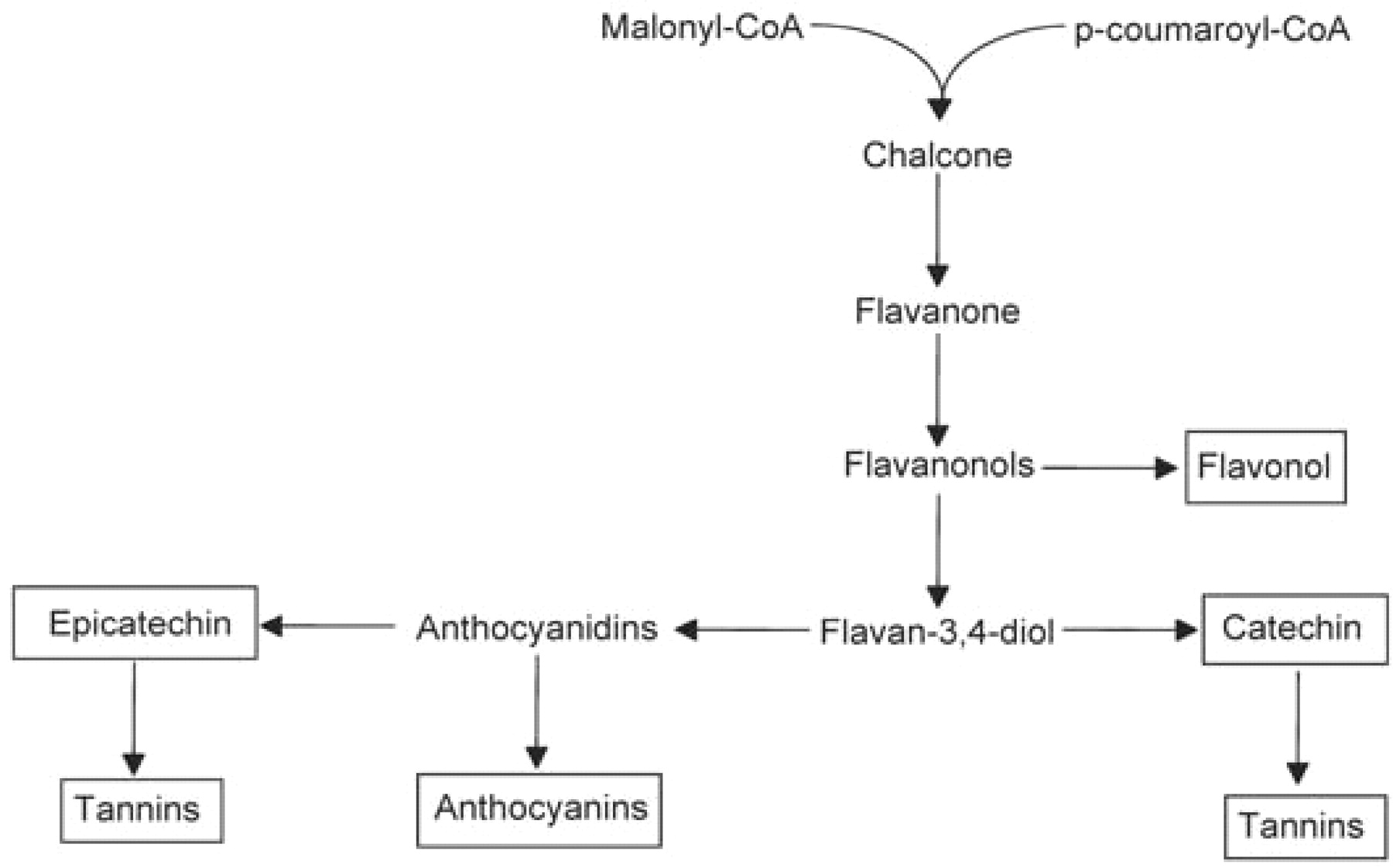
The biosynthetic relationship among the flavonoids that accumulate in various grape berry tissues is shown in Figure 8⇓, and the classes that accumulate are outlined by rectangles. The biosynthetic sequence leading to flavonoids begins at the chalcone synthase reaction where p-coumaroyl-CoA combines with three malonyl-CoAs to give a chalcone and CO2 (Figure 9⇓). It is interesting that grape berries do not appear to accumulate pelargonidin, the anthocyanin having only a single hydroxyl group on the B ring. If the CoA ester used by chalcone synthase is that of caffeic acid, then the chalcone product would have the substitution pattern on the B ring that corresponds to that found in catechin, epicatechin, and cyanidin (that is, an ortho dihydroxy configuration on the B ring). However, if the chalcone synthase reaction in grape uses predominantly p-coumaroyl-CoA as is typically shown, then the grape must also have very efficient flavonoid 3′ hydroxylases and flavonoid 3′5′ hydroxylases to account for the predominance of the dihydroxylated and trihydroxylated compounds found in the berry. The presence of kaempferol and dihydrokaempferol along with their 3-substituted glycosides supports the idea that at least some of the chalcone synthase utilizes p-coumaroyl CoA . In any case, hydroxylation of the B ring to give the dihydroxylated and trihydroxylated derivatives is thought to occur at the level of the flavonone or flavonol (Boss et al. 1996).
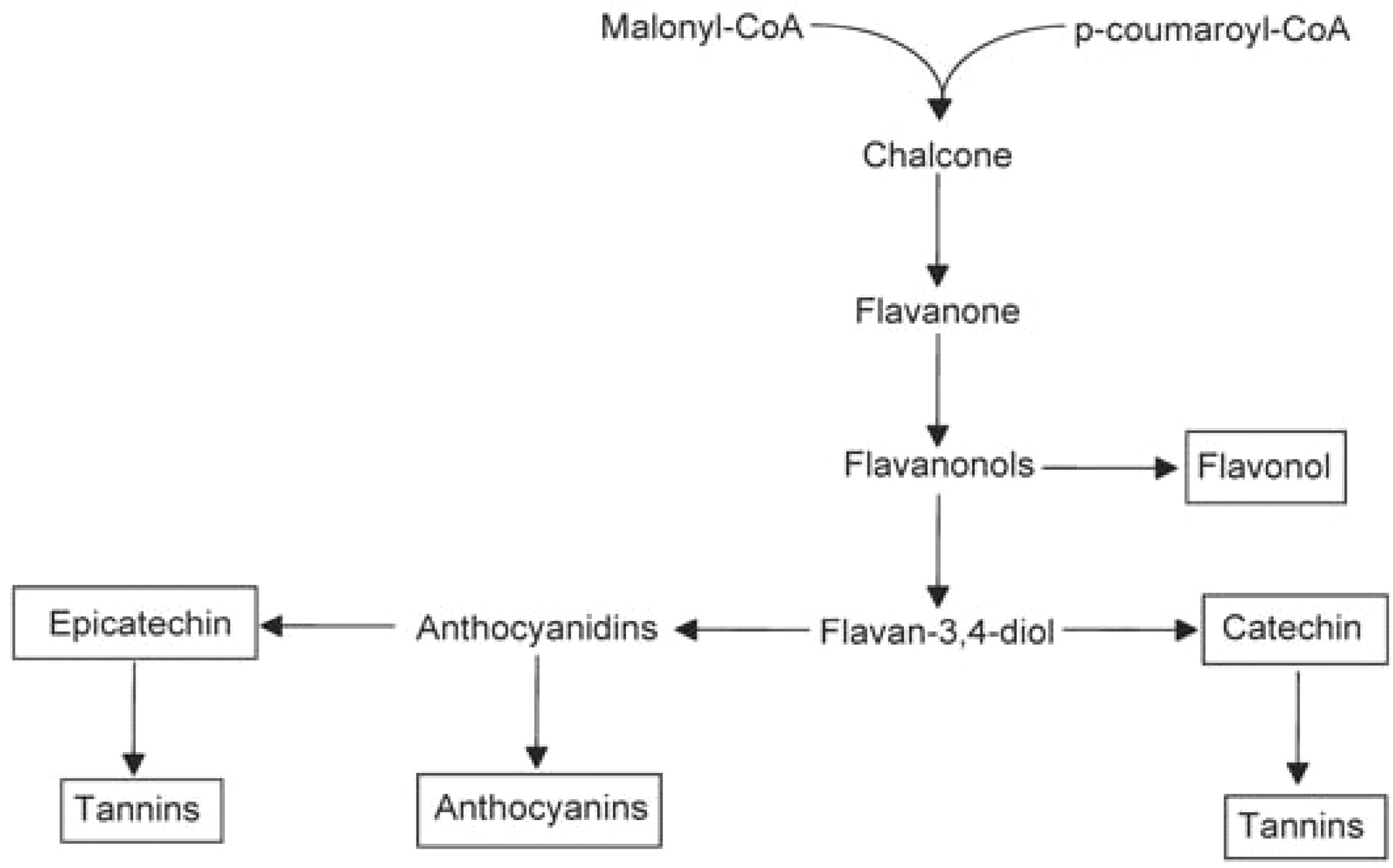
The biochemical pathway to the major flavonoids found in grape berries. Compounds that accumulate in the fruit are outlined by rectangles and the intermediates are present at very low levels.

Biosynthesis of the flavan-3,4-diol from caffeoyl-CoA and malonyl-CoA. CHS, chalcone synthase; CHI, chalcone isomerase; F3H, flavanone-3-hydroxylase; DFR, dihydroflavonol-4-reductase.
Anthocyanidins: Intermediates in Tannin Biosynthesis
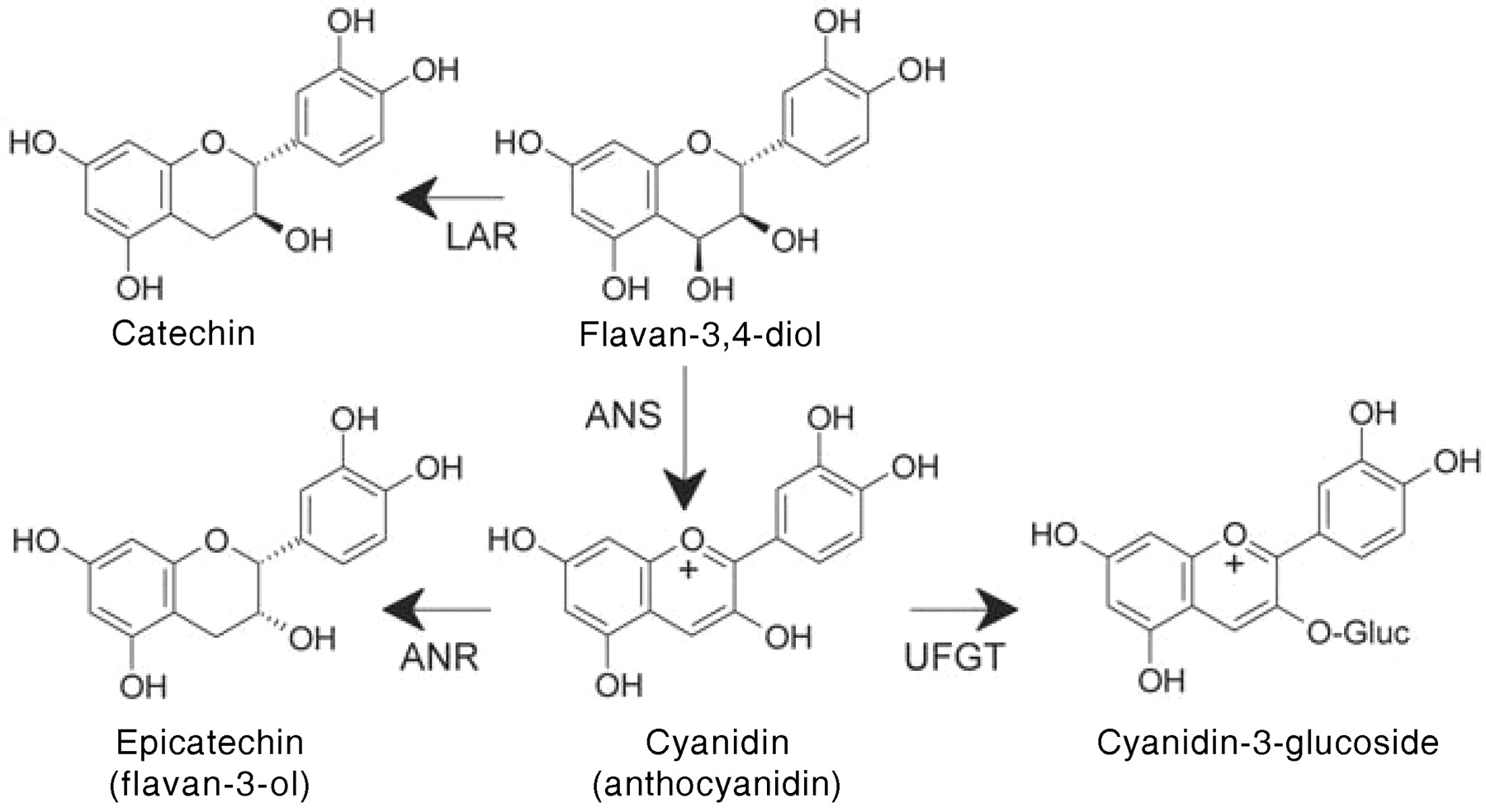
Recent work on tannin biosynthesis in the seed coat of Arabidopsis has revealed an unexpected relationship between anthocyanins and tannins (Xie et al. 2003). As described above, catechin is a 2,3-trans-flavan-3-ol whereas epicatechin is 2,3-cis. This stereochemistry is established at the chalcone isomerase reaction. This means that the flavan-3,4-diol produced from the flavanonol by dihydroflavanol reductase already has the 2,3,-trans configuration and can be converted to catechin directly by a leucoanthocyanidin reductase (LAR) (Figure 9⇑). An enzyme capable of catechin production from the 3,4,-diol was reported (Stafford and Lester 1984), but no enzyme capable of producing epicatechin (2,3,-cis) from the 2,3,-trans flavan-3,4-diol has ever been described. In 2003, Richard Dixon and colleagues showed that epicatechin is produced from cyanidin by the action of an enzyme (anthocyanidin reductase: ANR) that they described for the first time (Xie et al. 2003). Anthocyanidin reductase converts cyanidin and delphinidin to epicatechin and epigallocatechin, respectively. Since the flavan-3,4-diol looses its chirality in conversion to the corresponding anthocyanidin, biosynthesis of epicatechin proceeds through achiral anthocyanidin rather than by action of an epimerase and reductase on the 3,4-diol (Figure 10⇓). Epicatechin is the predominant extension unit of tannins in grape hypodermal cells and seeds, suggesting that cyanidin plays an important role as an intermediate in tannin biosynthesis in grape berries. In skin tissue when epigallocatechin is found as a tannin subunit, cyanidin and delphinidin would both be important intermediates in tannin biosynthesis. Bogs et al. (2005) found that in grape the ANR enzyme is encoded by a single gene whereas LAR is encoded by two closely related genes. The ANR gene was expressed in the skin and seeds of grape berries until the onset of ripening, and the two LAR genes showed different patterns of expression in skin and seeds. Timing and expression of ANR and LAR genes was consistent with the accumulation of proanthocyanidins in the berry, suggesting that they are responsible for determining tannin composition of the fruit during ripening. Several studies have shown the importance of UDP glucose-flavonoid 3-O-glucosyl transferase (UFGT) in anthocyanin accumulation in grape, and recognition of the aglycones as important intermediates in tannin production raises important questions about control and coordination of tannin and anthocyanin biosynthesis in grape tissues (Boss et al. 1996, Downey et al. 2004).

Formation of catechin, epicatechin, anthocyanidin (cyanidin), cyanidin-3-glucoside. LAR, leucoanthocyanidin reductase; ANS, anthocyanidin synthase; ANR, anthocyanidin reductase; UFGT, UDP glucose-flavonoid 3-O-glucosyl transferase.
Changes in the Phenolic Composition of Red Grape Berries during Ripening
The skin and seeds have features in common with regard to phenolics and ripening, but they also have their own unique characteristics. For example, both accumulate tannins as the most abundant class of phenolics at harvest, but anthocyanins only accumulate in the hypodermal cells of the skin. Enologically, the total amount of berry phenolics in each class at harvest is certainly important, but biologically it is useful to discuss the phenolic development of skin, mesocarp, and seed coat separately. In each case we will consider the phenolics of a particular tissue in order of their abundance.
Hypodermal cells of the skin.
Changes in tannin level in Cabernet Sauvignon fruit on a per berry basis during ripening are shown in Figure 11⇓, in which data for two consecutive years was taken from the same set of vines (Harbertson et al. 2002). Results show that skin tannins changed very little from veraison to harvest. Thus, tannins in the hypodermal tissue seem to be made very early in berry development and change very little from veraison to harvest on a per berry basis. Of course the concentration of skin tannin (mg/g fresh weight or mg/L) declines during ripening in proportion to berry growth during phase III. The values in Figure 11⇓ only reflect quantitative changes that take place and do not address qualitative changes (such as mDP) that might occur from veraison to harvest. Kennedy et al. (2001) showed that the mDP of skin tannins increased from 7.3 subunits in green berries of Shiraz three weeks after fruit set to 11.3 in red berries at veraison. By commercial harvest the mDP of skin proanthocyanidins in the Shiraz fruit reached 27. The situation with seed tannins is less clear: mDP determinations by thiolysis showing a decline from 8.29 subunits at veraison to 5.63 during ripening, whereas estimates made by HPLC predicted an mDP between 11 and 15 subunits as the fruit ripened (Kennedy et al. 2000a,b). These qualitative changes are probably very important for how tannins are perceived in the resulting wine matrix, but so far we have only mDP and degree of galloylation to characterize qualitative changes analytically. Nevertheless, both of these influence perception of tannin in wine, with mDP being the more important of the two (Vidal et al. 2003).

Changes in skin and seed tannin in Cabernet Sauvignon berries during ripening. Tannin is expressed in catechin equivalents (CE) on a per berry basis, 1998 and 1999 (Harbertson et al. 2002).
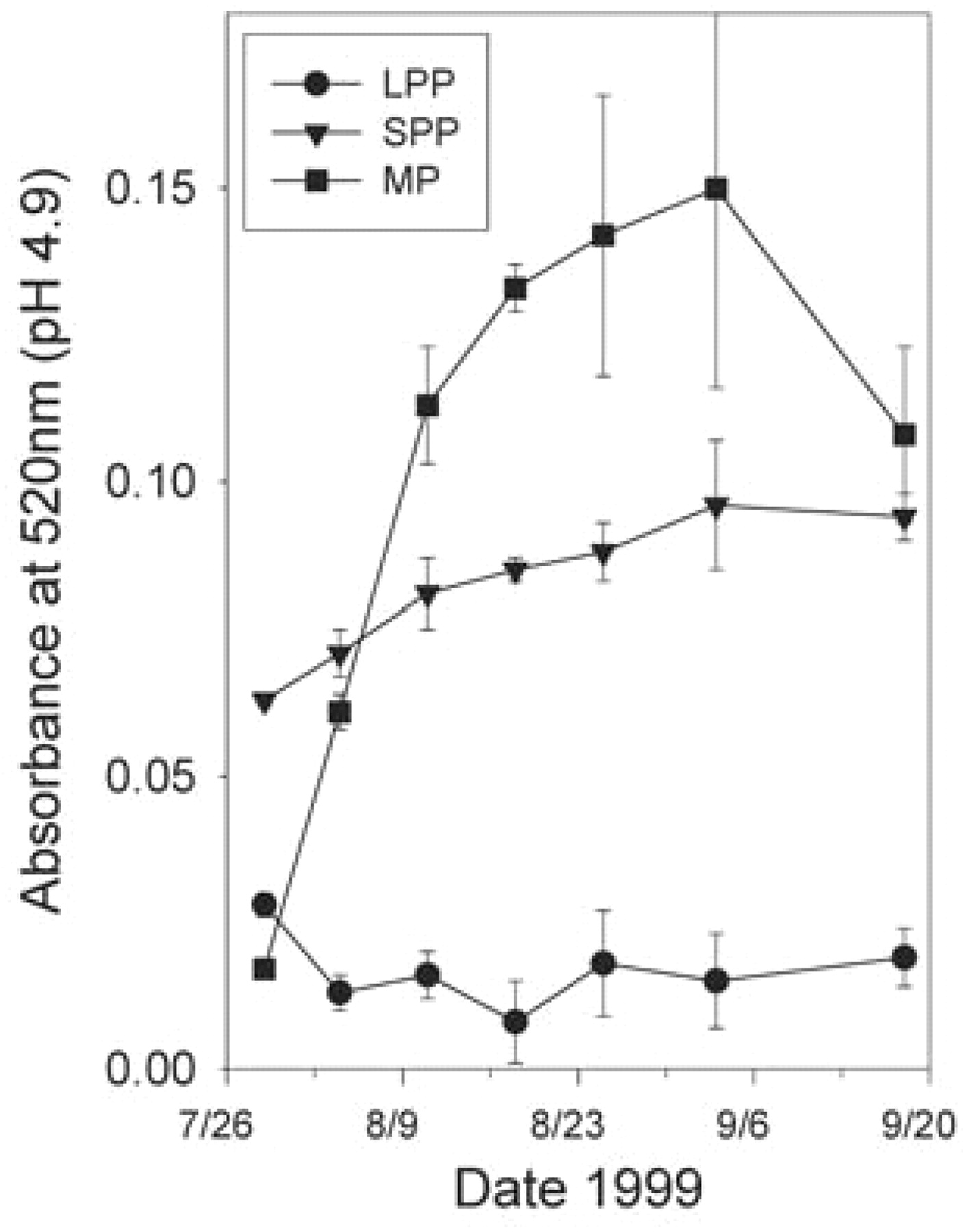
In addition to sugar increase, accumulation of anthocyanins in the skin of red varieties beginning at veraison must be one of the most commonly recognized features of berry ripening. Accumulation of total monomeric anthocyanins in the skins of Syrah berries beginning at veraison is shown in Figure 12⇓, which exhibits the typical anthocyanin accumulation curve and shows that the berry may contain polymeric pigments (Harbertson et al. 2003). One study used HPLC to quantify 15 monomeric anthocyanins in Shiraz berries during ripening (Boss et al. 1996). Malvidin glucosides dominated the accumulation curves, illustrating a common feature of many phenolic classes in grape berries: they are frequently dominated by one member of the class. So in the case of anthocyanins in Shiraz, malvidin-3-glucoside and its acylated derivatives predominate compared with the other four members of the class.
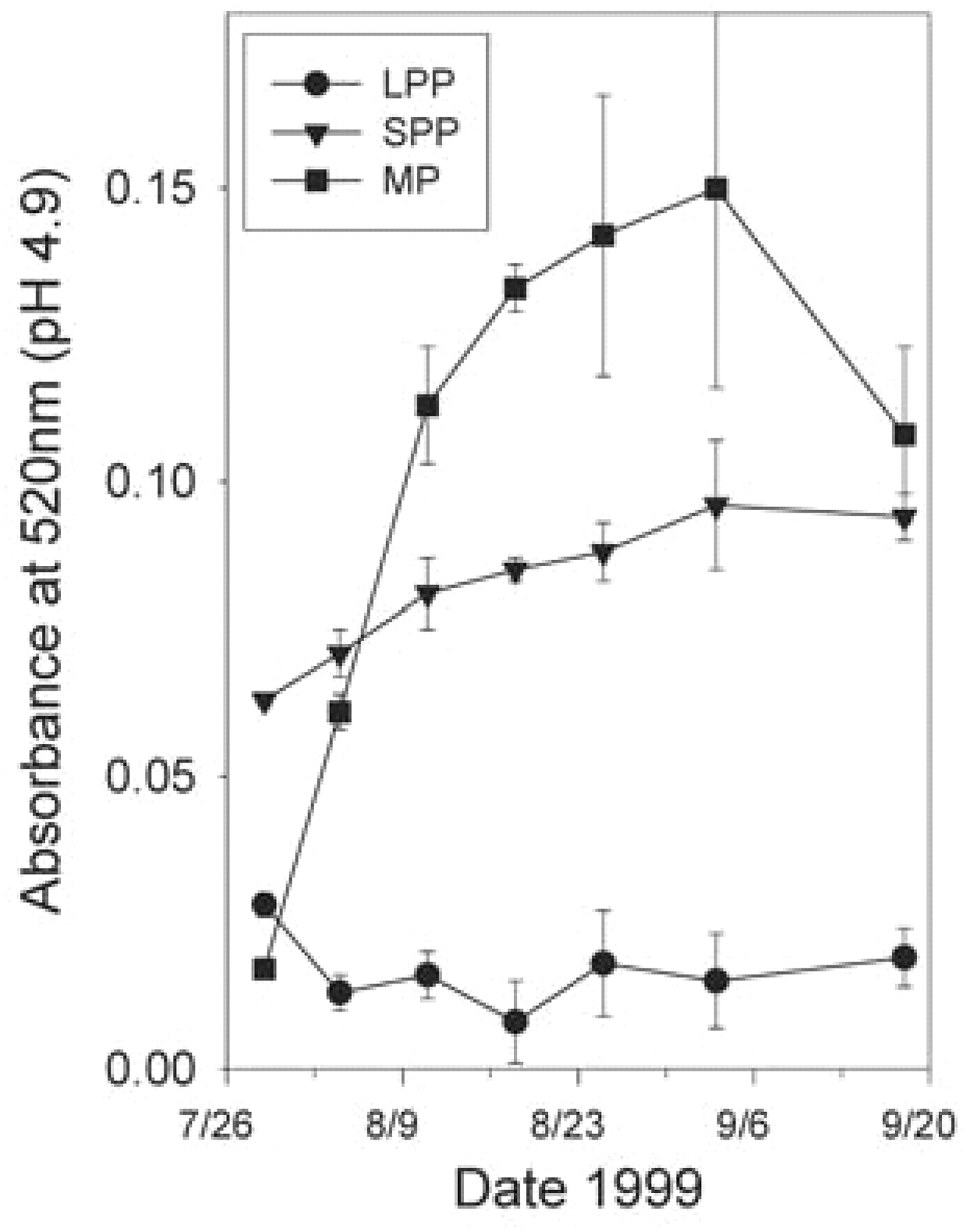
Changes in monomeric pigments (MP), large polymeric pigments (LPP), and small polymeric pigments (SPP) in Syrah berries during ripening (Harbertson et al. 2003).
The relative abundance of different phenolic classes in hypodermal cells was nicely illustrated in a study of flavonoids in the skin of a Vitis hybrid (DeChaunac) (Moskowitz and Hrazdina 1981). The authors measured anthocyanins, hydroxycinnamates, and flavonols in hypodermal cells and found that the anthocyanins were present at 2.88 mmol/g, of which 42% was represented by malvidin-3-di-glucoside, whereas the other four anthocyanins combined comprised the remaining 58%. Hydroxycinnamate esters were present at 0.98 mmol/g, only one-third the concentration of anthocyanins, and 77% of the hydroxycinnamates was caftaric acid, the remainder being coutaric and fertaric. The flavonol glycosides combined were 0.19 mmol/g, barely one-fifth the level of the hydroxycinnamates and only one-fifteenth compared to anthocyanins. Of the total flavonol glycosides, 93% was quercetin glycosides with only traces of the other flavonols shown in Figure 6⇑. This work illustrates the relative abundance of different phenolic classes in skin as well as the tendency for one member of the class to dominate the composition.
One final feature of phenolic phenomenology is the very uneven distribution of anthocyanin levels among different hypodermal cells of the skin. Hrazdina and Moskowitz (1982) reported that protoplasts prepared from skin of DeChaunac ranged from very light pink to very dark red, which was also noted in the study described above (Moskowitz and Hrazdina 1981). This variability of anthocyanin levels among different hypodermal cells suggests that they accumulate different amounts of anthocyanins, and it forces us to consider how this might be controlled at the level of individual cells when evaluating the biosynthetic schemes outlined in Figures 7⇑ and 8⇑.
Mesocarp cells of the pulp.
In most vinifera varieties the phenolic composition of mesocarp tissue is dominated by hydroxycinnamates. As described above, the hypodermal cells also contain hydroxycinnamates, but because of the larger contribution of the mesocarp to the total mass of the berry, changes in berry hydroxycinnamates during ripening usually reflect what occurs in mesocarp tissue.
Romeyer et al. (1983) described changes that occurred in the hydroxycinnamates during berry development and ripening. On a per berry basis total hydroxycinnamates showed a peak prior to veraison, then a decline leading to a constant amount (per berry) as the fruit ripened. This pattern of accumulation and decline for hydroxycinnamates is typical of several phenolic classes (for example, Figure 11⇑, and the flavan-3-ols described below). In each case it is not clear what becomes of the phenolics that disappear on a per berry basis during ripening. It may reflect catabolism of the compounds or their utilization in the biosynthesis of other phenolic classes, or both. In four varieties studied by Romeyer et al. (1983), caftaric acid was the predominant hydroxycinnamate followed by coutaric acid. Fertaric acid was only detected in two of the four varieties, and in those two it fell below detectable levels during ripening. Thus in ripe fruit the most abundant hydroxycinnamate was caftaric followed by coutaric.
A survey of hydroxycinnamates in ripe fruit showed that the level of hydroxycinnamates in the juice of different vinifera varieties was highly variable, ranging from 16 to 430 mg/L in the 37 vinifera varieties studied, with an average of 145 mg/L (Singleton et al. 1986). Cheynier et al. (1989) also found a range of hydroxycinnamate levels among white grape varieties studied. Thus, while the pattern of hydroxycinnamate accumulation and disappearance during ripening may be a general feature among varieties of Vitis vinifera, the amount present in the fruit of different varieties at harvest may represent a wide range. Unlike tannin and anthocyanins, little work has been done to determine how hydroxycinnamates vary in a single variety grown in different locations or in a single location using different viticultural practices.
Seeds.
The soluble phenolics of seed are mostly tannins, with lesser amounts of the monomeric flavan-3-ols catechin and epicatechin. The general decline in seed tannins during ripening as shown in Figure 11⇑ for Cabernet Sauvignon has been observed using different analytical methods. Ristic and Iland (2005) showed that this decline accompanies color changes that occur in seeds during ripening. Kennedy et al. (2000a, b) have suggested that this seed browning represents oxidation of tannins during ripening, but the chemical structures resulting from the presumed oxidation have yet to be described. Downey et al. (2003) have shown that the decline in seed tannin during ripening may reflect their covalent attachment to the insoluble matrix of the seed. They found that at the ripe stage up to 30% of the seed tannin extension subunits may be covalently bound and thus unavailable for extraction during winemaking.
It is interesting to note the difference in the pattern of tannin decline that occurred in seeds from the same vines in two consecutive years (Figure 11⇑). In 1998 the decline in seed tannin occurred soon after veraison and then showed no change in the four weeks prior to harvest. In the subsequent year, the decline occurred more slowly and only reached the low level of the previous year at time of harvest. It is unknown whether or not the pattern of tannin decline during ripening has any significance for wine-making (that is, influence on tannin extraction from seeds).
As pointed out above, tannins are a diverse set of compounds ranging in size from dimers to larger polymers, but they are generally considered as a single class because of the difficulty in separating individual chemical species. Nevertheless, one study reported how the family of B procyanidin dimers changed during ripening in seeds of four different varieties (Romeyer et al. 1986). The pattern of dimer accumulation appeared to be quite different in each of the four varieties that were studied.
The same study reported changes in catechin and epicatechin levels in seeds during ripening (Romeyer et al. 1986). Whereas the changes in B tannins appeared different in each of the four varieties, monomeric flavan-3-ols showed a very similar pattern during development. Each exhibited a maximum in both catechin and epicatechin levels in seeds just after veraison followed by a sharp decline as the fruit continued to ripen. The decline in seed catechin and epicatechin occurs concomitantly with seed browning, and the amount of the flavan-3-ols remaining in seeds at harvest appears to depend on the variety (Romeyer et al. 1986).
Summary and Conclusions
The berry structure and phenolic composition shown in Figure 1⇑ is useful for students in an introductory wine course but lacks sufficient resolution for winemakers faced with the task of optimizing extraction of these compounds during red wine fermentations. Recognizing the different phenolic composition of hypodermal cells versus mesocarp cells alone helps to explain why hydroxycinnamates are the most abundant phenolics in white juices and wines. However, unless we also consider the physical characteristics of the cells that make up the berry tissues to include the cell size and cell wall thickness, it is difficult to explain features of tannin and pigment extraction observed during red winemaking. Even within a single tissue type the phenolic composition can vary among adjacent cells, the best example perhaps being the different quantities of anthocyanin in different hypodermal cells. Thus, information about phenolic changes in grape berries during ripening must be organized and understood at the level of different tissues at a minimum and ultimately at the level of individual cell types within a tissue.
The biochemical pathways that plants have evolved to produce phenolic compounds from primary photosynthetic products have been described over the past 70 years. Knowledge of the fundamental details is still emerging, as illustrated by the recent discovery that anthocyanidins are obligatory intermediates in epicatechin and epigallocatechin biosynthesis. There are several critical features of phenolic biosynthesis that remain unknown; perhaps most conspicuous is the question of how the interflavan bonds of tannins are formed. In surveying research on polyphenol biosynthesis in a wide range of plants, it is clear that they share a common set of enzymes, but it is also clear that the way the pathways are regulated can be remarkably different among different species, and even among varieties of the same species. Thus, the biochemical networks responsible for the production of these compounds in grapes are reasonably well understood, but how the networks are regulated and coordinated in different varieties, tissues, and environments remains problematic.
Footnotes
Acknowledgments: Work from the author’s laboratory (Figures 11⇑ and 12⇑) was supported by the American Vineyard Foundation, the Viticulture Consortium, and the California Competitive Grant Program for Research in Viticulture and Enology. I thank the ASEV for the opportunity to present this work in the phenolics symposium and the reviewers for making many helpful suggestions for revising the manuscript.
This article was originally presented at the ASEV 56th Annual Meeting Phenolics Symposium, 20–21 June 2005, Seattle, WA. All phenolics symposium articles were peer reviewed by two fellow presenters, and James Harbertson, Mark Downey, and Sara Spayd served as technical editors of the articles.
From the ASEV 2005 Phenolics Symposium
- Copyright © 2006 by the American Society for Enology and Viticulture
Literature Cited
Vol 57 Issue 3








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
More from this TOC section
Similar Articles
