


Abstract
Canopy management is vital for quality winegrape production. During the 2012 and 2013 growing seasons, the timing of fruit-zone leaf removal (FZLR) was evaluated in two commercial vineyard blocks (Vitis vinifera Riesling and Sauvignon blanc) located north of Prosser, WA. Three different timings of manual FZLR were evaluated with a no-removal control. Leaf removal consisted of complete removal of all leaves and lateral shoots in the fruit zone on both sides of the canopy at prebloom, bloom, and four weeks postbloom. Each vine received the same treatment in both years. No negative implications were observed in total fruit set in either year. When leaf removal was performed, regardless of timing, the fruit zone of the canopy had less lateral shoot development and canopy refill than the control. Leaf removal also improved spray coverage in the fruit zone in Riesling, but the effect was related to the timing of leaf removal relative to the timing of the spray. In 2013, prebloom leaf removal significantly reduced Botrytis bunch rot severity in Sauvignon blanc (p = 0.01) below that of the control and four weeks postbloom leaf removal treatments. In 2013, prebloom leaf removal in Riesling increased terpene concentrations in the harvested juice (p = 0.03). In 2012, postbloom leaf removal in Riesling reduced concentrations of acids below those of the prebloom treatment (p = 0.04) in juice.
Fruit-zone leaf removal (FZLR) is a popular canopy management practice employed in winegrape (Vitis vinifera) growing regions around the world. This practice is typically carried out between fruit set and veraison (Diago et al. 2010, Percival et al. 1994a). In recent years, the practice of FZLR before fruit set has become more popular. Pioneering work from around the world has focused on the impact of timing and degree of FZLR on overall vine growth and development, cluster disease severity, and fruit composition (Bledsoe et al. 1988, Hunter et al. 1995, Lee and Skinkis 2013, Poni et al. 2006, Sabbatini and Howell 2010, Zoecklein et al. 1992).
Removing leaves in the fruit zone at prebloom can reduce the total canopy leaf area by over 50% (Poni et al. 2008), potentially resulting in insufficient carbohydrates for plant use during bloom and fruit set (Kliewer and Antcliff 1970). This may explain why some studies reported reduced fruit set and yield when FZLR was implemented prior to and during bloom (Palliotti et al. 2012, Percival et al. 1994b, Poni et al. 2009, Sabbatini and Howell 2010, Tardaguila et al. 2008). While reduced fruit set may be desired in locations where crop management techniques are legally restricted or where loose cluster architecture is desired, it is not always a universal production goal. In eastern Washington state, reaching contract-specific yields while maintaining quality is paramount. Reduced fruit set may negatively impact this goal and has resulted in caution regarding adopting early FZLR. Looser clusters as a means to reduce the risk of Botrytis bunch rot is not always a primary goal, as the climate of the region is not conducive to disease outbreaks on an annual basis. Washington state growers focus on ways to improve pest management programs (e.g., improved coverage of powdery mildew fungicides) and alter components like fruit microclimate that influence wine style and composition.
Mixed results are reported on the impacts of early FZLR on juice composition at harvest (Percival et al. 1994b, Poni et al. 2006, Staff et al. 1997, Tardaguila et al. 2008). Prebloom leaf removal in Barbera resulted in fruit with lower titratable acidity (TA) at harvest relative to an unthinned control (Poni et al. 2009). FZLR around fruit set in both Riesling and Chardonnay did not affect juice pH or TA (Zoecklein et al. 1992). In general, however, most authors reported either unchanged or increased harvest TA. Several studies reported reduced powdery mildew (Erysiphe necator) and Botrytis bunch rot (Botrytis cinerea) disease severity (Diago et al. 2010, Percival et al. 1994b, Sabbatini and Howell 2010, Staff et al. 1997). Both of these diseases can negatively affect overall juice composition at harvest.
The objectives of this study were to evaluate the timing of complete, early FZLR on canopy development, fruit set, spray coverage, and fruit composition in Riesling and Sauvignon blanc to optimize both horticultural and disease management attributes of this cultural practice under the arid conditions in eastern Washington state.
Materials and Methods
Vineyard description.
This trial was conducted in a commercial vineyard located northeast of Prosser, WA (lat. 46°15′36″N; long. 119°43′12″W) from spring 2012 to winter 2014, or two full growing seasons. Site soils are Warden-Silt loam, are well drained, and have a high water-holding capacity. One block each of own-rooted Riesling and Sauvignon blanc were used. Blocks were planted in 2007, with north-south row orientation. Planting distances were 2.7 m × 1.8 m (rows × vines) in Riesling and 2.7 m × 2.1 m in Sauvignon blanc. All vines were pruned to 14 three-bud spurs. Water was applied through drip irrigation. In both years, an initial irrigation application in early to mid-May was followed by subsequent applications after fruit set. Post-fruit set irrigation was designed as a regulated deficit of ~80% evapotranspiration (ETo) and irrigation sets were weekly to biweekly depending on plant response. In both years, a final irrigation set was done in October to replenish the soil moisture profile prior to dormancy. The canopy was trained using a modification of the vertical shoot-positioning (VSP) system. The modification consisted of only training one-third of the shoots to an upright position using fixed catch wires. The remaining shoots were allowed to sprawl on either side of the canopy. This modification is common in eastern Washington to reduce excessive sun exposure on the fruit. Summer canopy hedging was performed in mid-July after all leaf removal treatments had been executed. At the time of mechanical hedging, canopies were 1.25 m high, and the top 8 to 12 cm of 40% of the shoots was removed during this process. Hedging was coupled with regulated deficit irrigation to reduce further canopy development. All other management practices (e.g., insect pest, disease, nutrient) were carried out per the vineyard’s standard production procedures.
Weather.
Weather data was collected using Washington State University’s AgWeatherNet system (http://weather.wsu.edu). The “WSU-Prosser” weather station is located ~1.6 km from the research site. Average daytime high and low temperatures and total precipitation data were recorded, and growing degree days (base of 10°C; 1 April to 31 October) and evapotranspiration (ETo) were calculated.
Leaf removal treatments.
Leaf removal was done manually. On each treatment date, all leaves and lateral shoots present at the time of treatment were removed from the base of each count shoot up to the distal cluster, which was typically present on node four or five in both varieties. All non-count shoots and cordon suckers were removed prior to implementing the first leaf removal treatment in both varieties. Each leaf removal treatment was applied at a key phenological development stage as defined by the BBCH scale (Lorenz et al. 1994). The leaf removal timings evaluated were: no leaf removal (control), prebloom (~BBCH 57), bloom (BBCH 65, when 50% of inflorescences were at 50% capfall), and four weeks postbloom (BBCH 75). Dates of leaf removal in Riesling were 23 May 2012 and 14 May 2013 for prebloom, 13 June 2012 and 5 June 2013 for bloom, and 11 July 2012 and 3 July 2013 for four weeks postbloom. Dates of leaf removal in Sauvignon blanc were 30 May 2012 and 20 May 2013 for prebloom, 15 June 2012 and 7 June 2013 for bloom, and 12 July 2012 and 3 July 2013 for four weeks postbloom.
Leaf removal treatments were replicated four times in a randomized complete block design. Each treatment was applied to 24 vines (eight vines per row, in three adjacent rows) in each replicate (block). The six center vines in the center row of each treatment replicate were used for data collection and observation, allowing a one-row buffer on either side of the treatment and a one-vine buffer within the center row. The same treatments were imposed on the same vines in both years.
Leaf area removed.
In 2013, ten shoots per treatment were arbitrarily collected from vines outside of the experimental design to estimate the approximate leaf area removed during each treatment application. This method was used because the participating grower had already contracted much of the fruit in the research location and did not want full-vine defoliation at that time of year. Shoots were collected ten or four days after prebloom leaf removal in Riesling or Sauvignon blanc, respectively. The delay in shoot collection was a result of delayed vineyard entry due to a combination of timing overlaps relating to pesticide reentry periods. Shoots were collected within two days of the bloom treatment and within one day of the four weeks postbloom leaf removal treatment in both varieties. To measure leaf area, each shoot was stripped of all leaves and lateral shoots. Individual leaf area was then estimated for each leaf by multiplying the length of the midvein by the width of the leaf at the widest part. To calculate leaf area removed, leaf area of leaves found within the fruiting zone was compared to total leaf area for each shoot. This was then extrapolated to the whole vine.
Summer lateral shoot development.
To evaluate the degree of canopy refill in the fruiting zone, the presence and length of summer lateral shoots that remained after leaf removal (i.e., that were not present at the time of leaf removal) were determined in the fruit zone on ten shoots in each treatment replicate. In 2012, summer lateral shoot presence and the length of those laterals arising between nodes one and four on each main shoot were recorded on 15 Aug. In 2013, summer lateral shoot presence and length of lateral shoots arising between nodes one and five were recorded on 10 Sept and 29 Aug for Riesling and Sauvignon blanc, respectively. Summer lateral shoot presence and length was rated categorically: (i) no lateral shoot present, (ii) lateral shoot ≤ 3.0 cm, (iii) lateral shoot between 3.1 and 15.0 cm, and (iv) lateral shoot >15.0 cm.
Spray coverage.
The impact of FZLR on fruit-zone spray coverage was evaluated in 2013. Spray coverage was assessed on 20 June for both varieties (bloom) and again on 30 July for Riesling and 1 Aug for Sauvignon blanc (just prior to the onset of veraison). In each treatment replicate, one water-sensitive card (Syngenta Crop Protection AG, Basel, Switzerland) was placed in the vine canopy. Cards were affixed to the node between the basal and secondary clusters using a clothespin, with the water-sensitive side facing the row middle (i.e., outside of the canopy). The cards were placed in the vineyard just prior to spraying and were removed promptly after drying (~2 to 3 hr). Coverage was estimated on each card using open-source ImageJ software that calculates pixel areas using color thresholds (Abramoff et al. 2004).
Disease and sunburn severity.
The incidence and severity of Botrytis bunch rot and the severity of sunburn were evaluated. Severity was visually rated as percent cluster surface area affected. Botrytis bunch rot was rated as clusters expressing symptoms of internal berry rot (i.e., brown discoloration of berries without the presence of fungal sporulation, but without the acetic acid odors present to distinguish it from sour rot) or as rot with associated fungal sporulation. Given the dry harvest conditions during the evaluation years, all Botrytis bunch rot occurred as a nonsporulating, internal berry rot. Ratings were completed on ten arbitrarily selected clusters within a treatment replicate. Evaluation of clusters in both years and varieties occurred early to mid-September (preharvest, BBCH 89). In both years and varieties, the dual-action powdery mildew-Botrytis bunch rot fungicide Inspire Super (Syngenta, Greensboro, NC; difeconazole + cyprodinil) was used during bloom; no additional Botryticide applications were made after bloom in 2012. In 2013, in Sauvignon blanc, an additional Inspire Super application was used at the start of veraison (1 Aug) and Elevate (Arysta Lifescience, Cary, NC; fenhexamid) was applied on 13 Aug.
Fruit set and berry weight.
Fruit set was evaluated in both years. In 2012, eight or seven basal clusters of Riesling or Sauvignon blanc, respectively, in each treatment replicate were used to calculate fruit set. In 2013, ten basal clusters per treatment replicate were used in both varieties. Calyptras were collected using handmade 15 cm × 12 cm fine mesh white nylon bags (also referred to as “tulle”) as previously described (Keller et al. 2001). Bags were affixed to the selected basal clusters at prebloom (BBCH 57) and were removed after the completion of bloom (BBCH 71). Caught calyptras were counted. After fruit set, the same clusters were destructively sampled to count total berries. Fruit set was calculated by dividing berries per cluster by calyptras per cluster.
Berry weights were evaluated in both years. In 2012, Sauvignon blanc and Riesling fruit was harvested on 10 and 18 Sept, respectively. In 2013, Sauvignon blanc was harvested on 29 Aug and Riesling on 16 Sept. Berry weights were based on 100 berries per treatment replicate in 2012 and on 60 berries per treatment replicate in 2013.
Fruit composition.
At harvest, the juice soluble solids (Brix), TA, and pH of fruit exposed to different timing of leaf removal were evaluated. Data was collected on 18 Sept 2012 and 16 Sept 2013 from Riesling and on 10 Sept 2012 and 5 Sept 2013 from Sauvignon blanc. All evaluations occurred within 10 days of commercial harvest. Three basal clusters per treatment replicate were used in 2012 and five basal clusters per treatment replicate were used in 2013. Within a treatment replicate, clusters were pooled and whole-cluster pressed. The resulting juice (~200 mL) was used for analysis. Of this juice, ~7.0 mL was used to measure soluble solids, TA, and pH, and 50.0 mL was stored at −18°C until transported for volatile and ammonia analysis. Juice soluble solids were measured using a digital refractometer (Quick-Brix 60; Mettler-Toledo, Schwerzenbach, Switzerland). Juice pH was measured using an electrode (InLab Versatile 413; Mettler-Toledo). Juice TA was measured and calculated as described (Iland et al. 2000).
Volatiles were analyzed from pressed grape juice using a modification of the methods of Francioli (1999) and Howard et al. (2005). Frozen juice samples described above were gradually thawed, then adjusted to pH 6.4 with 0.5% phosphoric acid (H3PO4) for acid hydrolysis of glycosides. Three mL juice was added to a 15.0 mL sample vial (Supelco, Bellefonte, PA) with 30% w/v sodium chloride and sealed with a silicon septa cover. The sample was magnetically stirred at 1200 rpm and heated to 50°C for two min prior to exposure of the solid-phase microextraction (SPME) fiber. A 60-μm polydimethylsiloxane divinylbenzene (PDMS/DVB) SPME fiber (Supelco) was exposed to the headspace of the sample for 60 min at 50°C with constant stirring. After extraction, fibers were desorbed in the injection port of a gas chromatography–mass spectrometry system (Hewlett Packard 5890II/5970; Agilent Technologies, Santa Clara, CA) with a (60 m × 0.32 μM) DB1 capillary column (Phenomenex, Torrance, CA). The mass spectrometer (ion source maintained at 250°C) used electron impact with electron energy of 70 eV. The SPME fiber was desorbed in the injection port for five min at 200°C using splitless injection. The capillary column was set at 33°C and held for five min before ramping to 50°C at a rate of 2°C/min. Mass spectral results were viewed using Chemstation G1803C software (Agilent Technologies). Compounds observed as chromatographic peaks were identified by comparing their mass spectra and retention times with monoterpene standards: linalool (≥95% GC, Fluka, Switzerland), geraniol (98% GC, Sigma-Aldrich, St. Louis, MO), cis-rose oxide (>99% GC, Fluka), nerol (≥90% GC, Fluka), 2- and 3-carene (95% GC, Sigma-Aldrich), α-terpineol (mix of isomers 95%, Sigma-Aldrich), and hexanal, nonanal, decanal, hexanoic acid, and decanoic acid (≥95%, Sigma-Aldrich). Identifications were made using published retention times and mass spectra of the Wiley and NIST library spectra database and the listed standards. There were four replicate extractions per treatment. Standard curves were generated by running compounds as mixtures from minimum detection to above the highest detected amounts in the study. The R2 of the standard curve was ≥85%.
The 50.0 mL juice samples used for volatile analysis were also used for ammonia analyses. Frozen samples were allowed to gradually thaw prior to total ammonia anlysis. A standard curve was created following manufacturer’s instructions (Ammonia Combination Electrode; Denver Instruments, Bohemia, NY) using 1, 5, 10, or 100 mg/L single-concentration standard solutions of ammonium (NH4+) and 0.2 mL allotments of 10 M NaOH. The standard curve was created twice. To evaluate ammonia in the juice samples, the same protocol for developing the standard curve was used, but with 10.0 mL juice rather than the standard and NaOH added in 0.4 mL allotments.
Skin and seed tannin and phenolic concentrations.
Tannin and phenolic concentrations in berry skins and seeds were evaluated in both years using 30 berries per treatment replicate. Berries, with pedicels attached, were removed from 10 or five (2012 or 2013, respectively) clusters at harvest. Berries were collected from various locations throughout each cluster, placed into plastic storage bags, and stored at −35°C (2012) or −80°C (2013) until analysis. Immediately prior to analysis, pedicels were removed from the frozen berries and the pool of 30 berries per treatment replicate was weighed. The skin of each berry was removed from the flesh by hand and seeds were extracted from the flesh and counted. Both skins and seeds were dried separately at room temperature for four hours and weighed. After drying, skins and seeds were placed into separate plastic vials and stored at −35°C (2012) or −80°C (2013) until tannin and phenolic extraction could be completed.
To extract seed tannins and phenolics, seeds were ground to a fine powder using liquid nitrogen in a sterile mortar and pestle. The powder was dissolved in 30 mL of 70% acetone. To extract skin tannins and phenolics, skins were transferred directly to a vial containing 30 mL of 70% acetone. Both skin and seed sample solutions were shaken for 12 to 24 hr at 100 rpm (SCILOGEX SK-330-Pro-Shaker, Berlin, CT). After agitation, samples were centrifuged at 5000 rpm for five min (Eppendorf 5804 R, Hamburg, Germany) and decanted into polyvac vials (PB3002-S; Mettler-Toledo). Samples were heated to 40°C and agitated under a vacuum at 300 rpm (Buchi Syncore Polyvap, Switzerland) until 13.0 to 15.0 mL of the sample remained. Post-evaporation, samples were weighed and transferred to plastic vials for storage at −80°C until total tannins and phenolics could be measured. Tannin and phenolic measurements were done as described (Hagerman and Butler 1978, Harbertson et al. 2003).
Statistical analysis.
Statistical analysis was completed using JMP statistical analysis software (JMP 9.0.0; SAS Institute Inc., Cary, NC). Data were tested for normality by the Shapiro-Wilk test and for homogeneity of variance by Levene’s test. No variables required transformation.
Data were subjected to analysis of variance (ANOVA), and means were separated using Tukey’s HSD at α ≤ 0.05.
Results
Weather.
In 2012 and 2013, the Yakima Valley American Viticultural Area accumulated 1468 and 1589 (°C) growing degree day units, respectively. The historical average for the area is 1406 (°C). Average monthly temperatures and average monthly minimum and maximum temperatures, monthly total solar radiation, evapotranspiration, and precipitation for May to September in 2012 and 2013 are reported (Table 1).
The monthly average maximum, average minimum, daily average temperatures, monthly total solar radiation, monthly total reference evapotranspiration, and monthly precipitation in 2012 and 2013 in Prosser, WA. The data are from AgWeatherNet (www.weather.wsu.edu); the weather station used was WSU-Prosser.
Assessment of leaf area removed.
Total leaf area (TLA) was assessed in 2013. The prebloom, bloom, and four weeks postbloom leaf removal treatments in Riesling removed 59.6, 49.7, and 22.4% of the TLA, respectively (Figure 1A). Riesling prebloom and bloom treatments removed a significantly higher proportion of the canopy at the time of treatment than postbloom leaf removal, as expected. Prebloom, bloom, and four weeks postbloom leaf removal in Sauvignon blanc removed 52.6, 35.0 and 18.3% of the TLA, respectively (Figure 1B). Sauvignon blanc prebloom treatment removed a significantly higher proportion of the canopy than the bloom and postbloom treatments, and the bloom treatment had significantly more canopy removed than the postbloom treatment. Additionally, with prebloom leaf removal assessments occurring several days after the treatment was implemented, it is expected that actual TLA removed may have been higher than recorded here.

Total leaf area removed as a percentage of the existing canopy at prebloom, bloom, and four weeks postbloom in 2013 for Vitis vinifera (A) Riesling and (B) Sauvignon blanc. Ten shoots per treatment were used to determine the total leaf area removed. Letters denote significant differences between treatments within each year, using Tukey’s HSD at α = 0.05. Bars denote standard error of the mean.
Summer lateral shoot development.
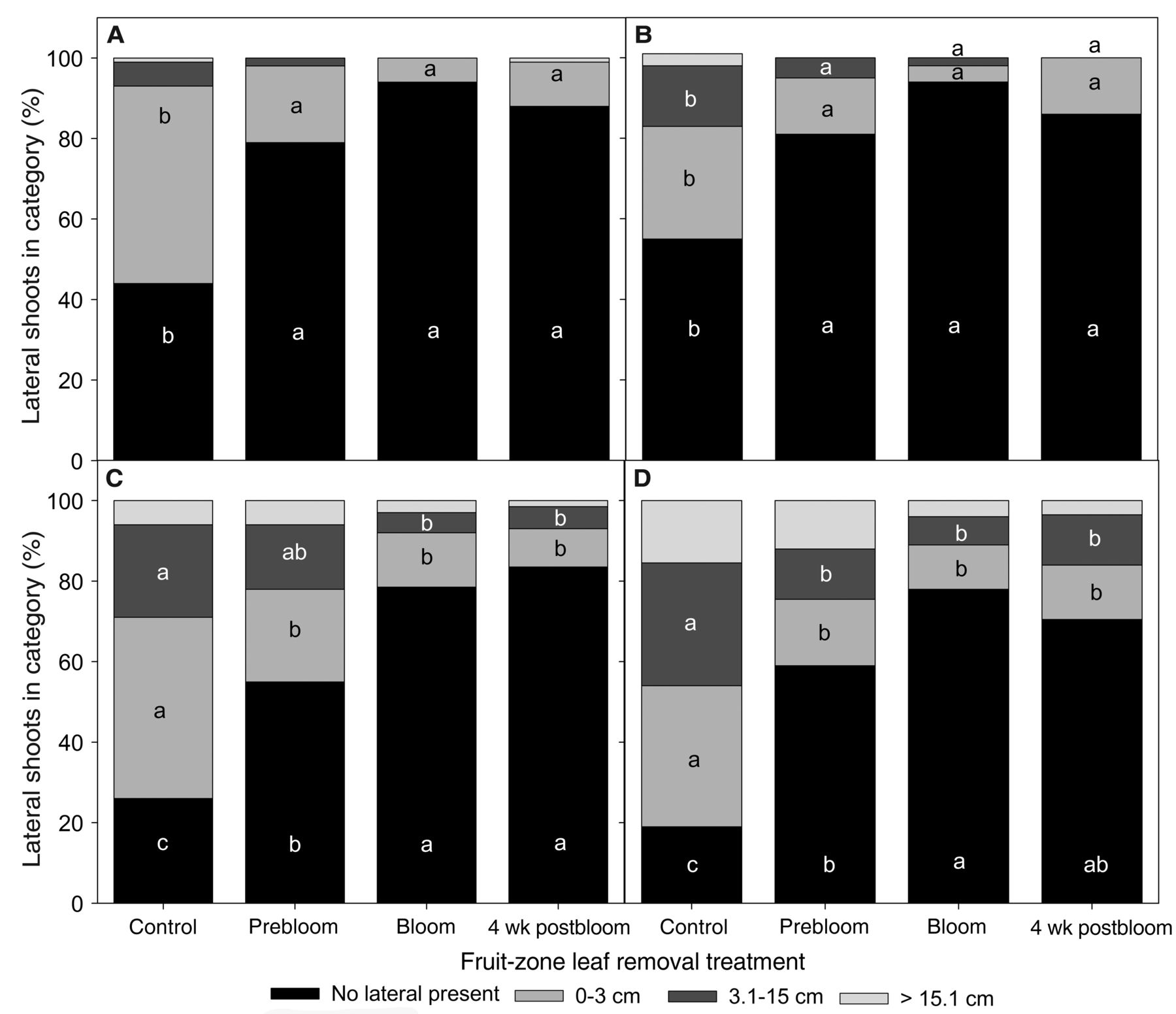
In both years and on both varieties and regardless of timing, leaf removal reduced the incidence of lateral shoot development in the fruit zone and the incidence of lateral shoots of between 0 and 3 cm (Figure 2). Leaf removal also reduced the incidence of lateral shoots of intermediate length in Sauvignon blanc below that of the control. There were also significant effects of leaf removal timing. In 2013, leaf removal at bloom was more effective at keeping the fruit zone free of lateral shoots than prebloom leaf removal in both Riesling and Sauvignon blanc (Figure 2C and 2D). In Riesling specifically, both bloom and postbloom leaf removal significantly lowered the incidence of lateral shoots of intermediate length in comparison to the control, whereas prebloom leaf removal did not (Figure 2C).

Lateral shoot development in the fruit zone (first four to five nodes on count shoots) of Vitis vinifera Riesling and Sauvignon blanc undergoing different fruit-zone leaf removal treatments. In 2012, lateral shoot growth was assessed and categorized on 15 Aug in (A) Riesling and (B) Sauvignon blanc. In 2013, lateral shoot growth was categorized on (C) 10 Sept in Riesling and (D) 29 Aug in Sauvignon blanc. Categories for shoot development included: (1) no laterals present; (2) laterals between 0 and 3 cm; (3) laterals greater than 3 cm but at or less than 15 cm; and (4) laterals greater than 15 cm. Letters denote significant differences between treatments within each year, using Tukey’s HSD at α = 0.05.
Spray coverage.
On both assessment dates in Riesling, bloom leaf removal improved spray coverage (Figure 3A). On 20 June, prebloom leaf removal provided significantly higher coverage. Unexpectedly, coverage in the four weeks postbloom leaf removal treatment, which had not occurred at the time of this spray application, was not different from the other leaf removal treatments, but as expected, it was also not different from the control. High variability in spray coverage results in late June may not be unusual, as canopies have not reached full size or density at this time. On 30 July, a time when all leaf removal treatments had been completed, four weeks postbloom leaf removal provided significantly higher coverage than the control, while prebloom leaf removal was not different than the control but was also not different from the other treatments (Figure 3A).

Spray coverage at bloom and preveraison between different fruit-zone leaf removal treatments in Vitis vinifera (A) Riesling and (B) Sauvignon blanc in 2013. Spray coverage near bloom in Riesling was assessed on 20 June; the preveraison spray coverage assessment was on 30 July. Spray coverage near bloom in Sauvignon blanc was assessed on 20 June; the preveraison spray coverage assessment was on 1 Aug. At the time of the bloom assessment, only the prebloom and bloom leaf removal treatments had been implemented. Water-sensitive cards were placed between basal and secondary clusters on count shoots just prior to spray application. Treatment means within an assessment date not connected by the same letter(s) denote significant differences using Tukey’s HSD at α = 0.05. Bars denote standard error of the mean.
There were no differences in spray coverage as a result of leaf removal timing on either assessment date in Sauvignon blanc (p = 0.75 and 0.08; Figure 3B). While the 20 June assessment date had a similar coverage pattern to Riesling, high level of coverage variability resulted in no statistical difference among treatment means. At the 1 Aug assessment, the control had the overall lowest average coverage (10.5%), while the leaf removal treatments ranged between three and four times more coverage than the control (34.0 to 41.4%) as in Riesling.
Disease and sunburn severity.
Botrytis bunch rot severity in Riesling was not influenced by leaf removal in either year (p = 0.28 and 0.15, respectively, for 2012 and 2013). Severity ratings in 2012 were 17.8, 29.5, 20.0, and 20.3%, for the control, prebloom, bloom, and postbloom leaf removal treatments, respectively. Severity ratings for Botrytis bunch rot in Riesling in 2013 were 19.5, 13.0, 6.8, and 6.3% for the control, prebloom, bloom, and postbloom leaf removal treatments, respectively.
Botrytis bunch rot severity in Sauvignon blanc was not influenced by leaf removal in 2012 (p = 0.43). Total disease severity was 19.8, 20.3, 19.0, and 22.5% for control, prebloom, bloom, and postbloom leaf removal, respectively. However, in 2013, the Sauvignon blanc prebloom leaf removal had lower disease severity (4.7%) than either the control (12.0%) or four weeks postbloom leaf removal (11.8%; p = 0.01 and 0.01, respectively). Bloom leaf removal had an intermediate level of disease (6.8%).
The timing of leaf removal did not influence sunburn severity in either year or variety. In 2012, sunburn severity for the control, prebloom, bloom, and four weeks postbloom treatments were 5, 7, 6, and 12% in Riesling (p = 0.36) and 5, 8, 14, and 16% in Sauvignon blanc (p = 0.09), respectively. In 2013, sunburn severity for the control, prebloom, bloom, and four weeks postbloom treatments were 7, 10, 8, and 20% in Riesling (p = 0.12) and 6, 8, 12, and 14% in Sauvignon blanc (p = 0.29), respectively.
Fruit set and berry weight.
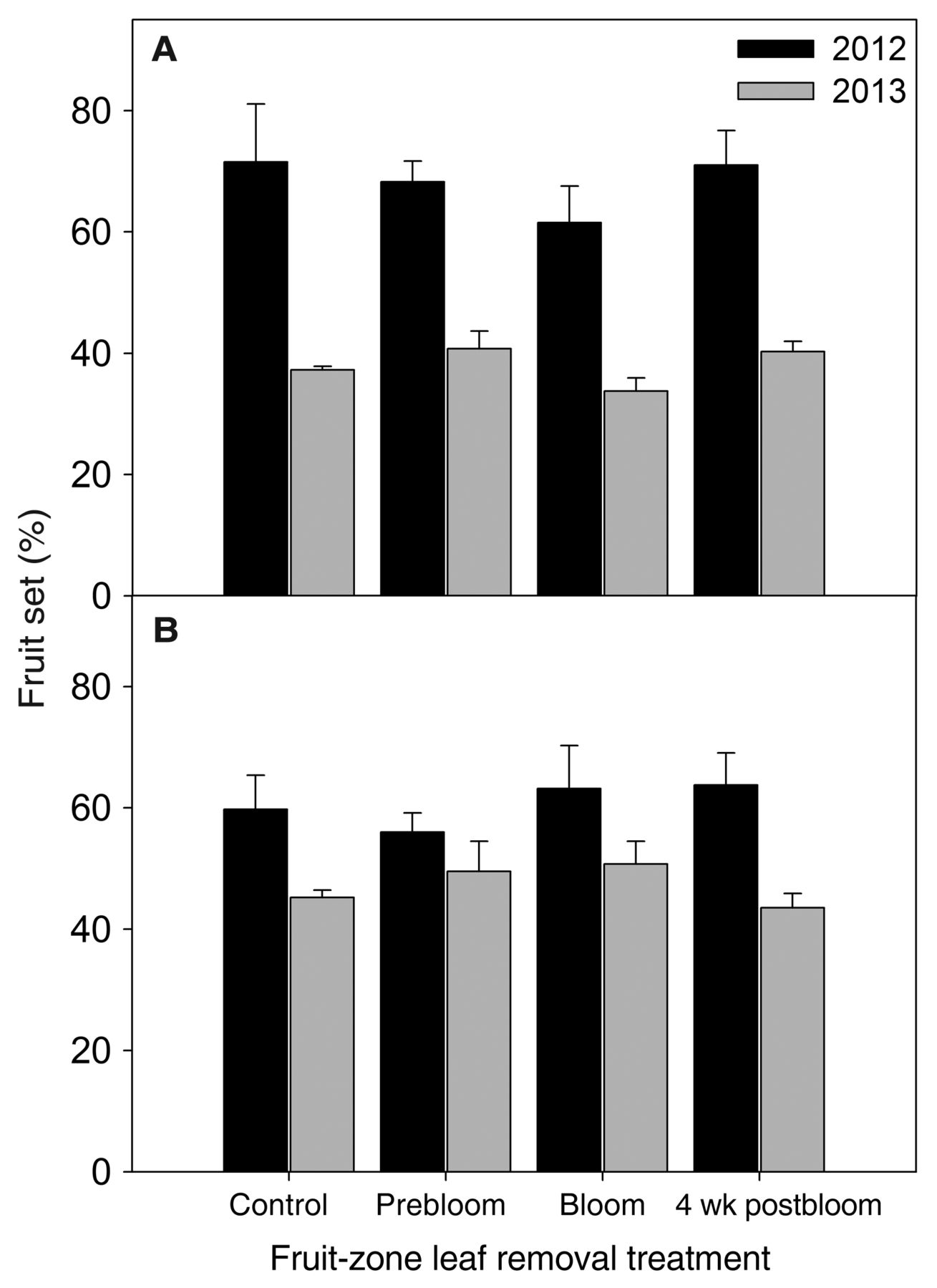
FZLR, regardless of timing, did not impact overall fruit set for Riesling (p = 0.60 and 0.05, respectively, in 2012 and 2013) or Sauvignon blanc (p = 0.65 and 0.30, respectively, in 2012 and 2013; Figure 4). Overall, 2012 had higher fruit set in both varieties than 2013, likely due to the more moderate temperatures and evaporative demands during bloom (Table 1).

Fruit set, expressed as percentage total flowers (estimated through calyptra counts) setting to berries for different fruit-zone leaf removal treatments in Vitis vinifera (A) Riesling and (B) Sauvignon blanc in 2012 and 2013. Bars denote standard error of the mean. No significant differences were seen among treatments in either variety or year.
Average berry weights for Riesling in 2012 were 1.27, 1.20, 1.23, and 1.23 g, respectively, for the control, prebloom, bloom, and four weeks postbloom treatments and were not significantly different from each other (p = 0.73). Average berry weights in 2013 were 1.24, 1.23, 1.20, and 1.22 g, respectively, for the control, prebloom, bloom, and four weeks postbloom treatments and were not different from each other (p = 0.89). Average berry weights for Sauvignon blanc in 2012 were 1.27, 1.22, 1.23, and 1.23 g, respectively, for the control, prebloom, bloom, and four weeks postbloom treatments and were not different from each other (p = 0.74). Average berry weights in 2013 were 1.18, 1.17, 1.16, and 1.10 g, respectively, for the control, prebloom, bloom, and four weeks postbloom treatments and were not different from each other (p = 0.62).
Fruit composition.
Leaf removal did not influence harvest soluble solids, TA, or pH in either variety in either year (Table 2). In addition, the timing of leaf removal did not influence total aromatic alcohols and terpenes in either variety in 2012 (Table 2) nor did it influence composition (all volatiles assessed) in Sauvignon blanc in 2013. However, bloom leaf removal in Riesling in 2012 did reduce total aromatic aldehyde concentrations (p = 0.05); specifically, hexanal was reduced (p = 0.03). Prebloom leaf removal in Riesling in 2012 increased total acid concentrations relative to four weeks postbloom (p = 0.04).
Juice composition as a function of leaf removal treatments in Vitis vinifera Riesling and Sauvignon blanc for the 2012 and 2013 vintages.
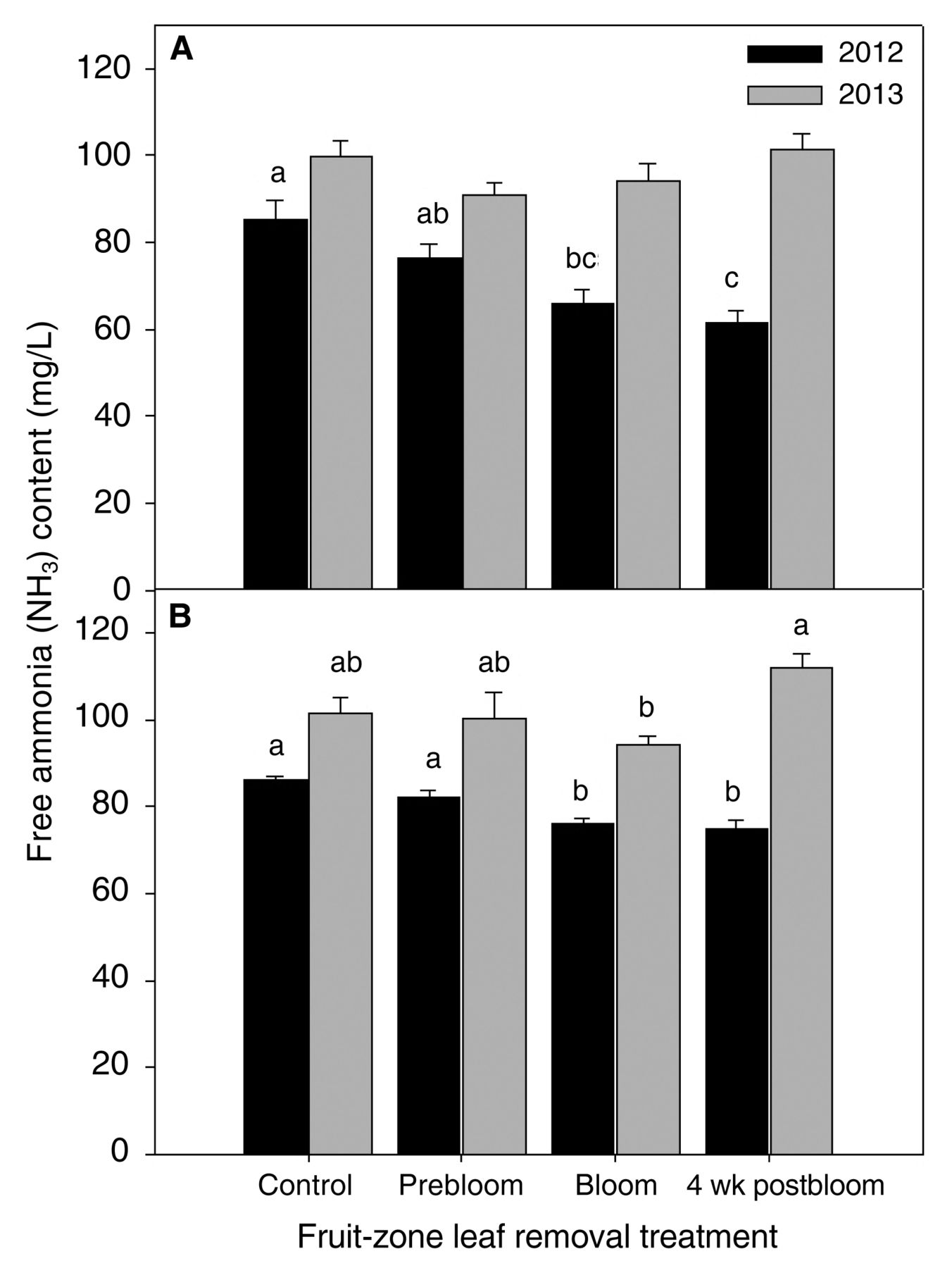
In 2013, the timing of leaf removal did not influence total aromatic aldehydes and acids in Riesling (Table 2). However, prebloom leaf removal did increase total terpene concentrations (p = 0.03); specifically, an increase in α-ionone relative to the control and bloom leaf removal (p = 0.003 and 0.05, respectively). Nerol oxide concentrations were significantly less in the control relative to prebloom and four weeks postbloom leaf removal (p = 0.02 and 0.02, respectively). The timing of leaf removal influenced total ammonia (NH3) in Riesling in 2012 (Figure 5). Bloom and four weeks postbloom leaf removal reduced ammonia relative to the control (p = 0.003 and 0.0006, respectively). The four weeks postbloom leaf removal also reduced ammonia relative to prebloom (p = 0.02). Leaf removal did not influence total ammonia in 2013.

Free ammonia (NH3) concentrations in juice from Vitis vinifera (A) Riesling and (B) Sauvignon blanc subjected to different fruit-zone leaf removal treatments in 2012 and 2013. Treatment means within a year and variety not connected by the same letter(s) denote significant differences using Tukey’s HSD at α = 0.05. Bars denote standard error of the mean.
Leaf removal did not influence aromatic volatiles in Sauvignon blanc in either year (Table 2). It did, however, influence total ammonia (Figure 5). In 2012, bloom and four weeks postbloom leaf removal decreased total ammonia relative to the control and prebloom leaf removal (p = 0.0002; Figure 5). In 2013, bloom leaf removal reduced total ammonia relative to four weeks postbloom leaf removal (p = 0.02). In both varieties, prebloom leaf removal was the only leaf removal timing that did not reduce total ammonia relative to the control.
Skin and seed tannin and phenolic content.
The timing of leaf removal in Riesling did not alter total skin or seed tannins or phenolics in either year (Table 3). In Sauvignon blanc, the timing of leaf removal influenced seed total phenolics in 2012, seed tannins in 2013, and skin tannins and phenolics in both years (Table 3). In 2012, prebloom and four weeks postbloom leaf removal increased total seed tannins relative to the control (p = 0.001 and 0.001, respectively), as well as skin phenolics relative to the control (p = 0.01 and 0.001, respectively). In 2013, bloom leaf removal had higher skin phenolic concentration, seed tannin concentration, and skin tannins relative to the control (p = 0.008, 0.005, and 0.005, respectively). Bloom leaf removal also resulted in higher skin tannins than four weeks postbloom leaf removal (p = 0.005).
Skin and seed tannins and total phenolics as a function of leaf removal treatments in Vitis vinifera Riesling and Sauvignon blanc for the 2012 and 2013 vintages.
Discussion
FZLR starting as early as prebloom did not affect key production parameters in Riesling and Sauvignon blanc negatively. Growers initially feared that complete leaf removal in the fruit zone prior to fruit set would reduce fruit set, because production goals in eastern Washington are aimed at optimal yield rather than a reduction in cluster compactness to combat diseases. This focus on yield rather than disease management is expected in a location where environmental conditions do not favor widespread incidence of various harvest rots (Table 1). However, the same conditions that reduce disease pressure are also often associated with increased sunburn and decreased fruit quality (Spayd et al. 2002). Thus, early FZLR and leaf removal on both sides of the canopy has not been widely adopted. In 2012 and 2013, which were “average” and “above average” in heat, neither reduced fruit set nor reduced fruit quality were seen as a result of FZLR, suggesting that early leaf removal on both sides of the canopy in a modified VSP system might be an appropriate cultural practice for eastern Washington grape growers.
The lack of affect on fruit set may stem from the environmental conditions in eastern Washington or from the severity of leaf removal used. The complete removal of leaves at four to five basal nodes used here, while more severe than current grower standards (i.e., leaf removal on the east or north sides of the canopy only), was likely not severe enough to induce significant changes in plant source-sink relationships under the typical growing conditions and season length seen in Washington (Table 1). In studies where the severity of leaf removal was higher, leaves from the medial area of the main shoot upward had a higher photosynthetic capacity and compensated for the loss of older basal leaves (Poni et al. 2006). Removing the first six basal leaves on shoots at prebloom resulted in higher net canopy CO2 exchange rates than in vines without FZLR, with the compensation peaking ~15 days postdefoliation (Poni et al. 2008). Coinciding with the increased CO2 exchange rates, carbohydrate concentration also increased.
While the degree of FZLR presented here is less severe than in other studies, it still improved spray coverage during the critical period of fruit susceptibility to diseases such as powdery mildew and Botrytis bunch rot, which occurs around bloom (Ficke et al. 2003, McClellan et al. 1973). In Riesling, leaf removal at prebloom and bloom allowed significantly higher spray coverage than that of the control or postbloom leaf removal. While spray coverage after veraison was not assessed in this study, leaf removal, regardless of timing, had significantly fewer instances of summer laterals in the fruit zone than the control (Figure 2C), which would improve air circulation and sunlight penetration, reducing the microclimate favorability for Botrytis bunch rot (English et al. 1990). Interestingly, in our study, there was only one variety-year combination with leaf removal that significantly reduced Botrytis bunch rot: prebloom leaf removal in 2013 in Sauvignon blanc. This also happened to be a variety and year without significant differences in spray coverage. Botrytis bunch rot incidence is low to nil in most years in eastern Washington due to the lack of conducive environmental conditions during fruit ripening. In this situation, during the bloom-time pesticide application for Botrytis bunch rot control, both prebloom and bloom leaf removal treatments had a more exposed fruit zone for reduced environmental favorability for B. cinerea colonization, despite a lack of difference in spray coverage. At the end of the season, however, the prebloom leaf removal timing still had a more open fruit zone than the bloom treatment (Figure 2D), potentially allowing improved coverage for the postveraison Botryticide applications made in that year. The reduced Botrytis severity seen in the prebloom leaf removal treatment in Sauvignon blanc may be due to improved coverage during those later applications (data not collected) or related to specific alterations in canopy microclimate at key times, thus reducing initial colonization by the fungus. The authors speculate that in years with more favorable environmental conditions for Botrytis bunch rot development during veraison, early FZLR would result in improved rot control. Additionally, a more severe application of FZLR at prebloom or bloom (i.e., to above the fruit zone) might reduce fruit set, resulting in looser clusters with a reduced risk for Botrytis bunch rot disease severity.
Past studies also demonstrated that removing the six basal leaves on shoots during rachis elongation (BBCH 57) resulted in strong lateral shoot growth (Kriedemann 1968, Reynolds and Wardle 1989). Canopy hedging, which results in loss of apical dominance, also promotes growth of lateral shoots. In certain environments, this may require additional vineyard passes to maintain an open fruit zone. In eastern Washington, more than one pass is not desired by growers due to the increased vineyard management costs. At the same time, a completely exposed fruit zone that might result in sunburned fruit is also a concern. As such, some lateral shoot growth in the fruit zone is desirable, provided the other benefits of leaf removal like reduced disease pressure and improved spray coverage remain optimal. Interestingly, in eastern Washington, grapevine canopy size is predominately controlled through the use of regulated deficit irrigation, which can limit the development of lateral shoots even when coupled with hedging. In our study, if lateral shoots were present at the time of leaf removal, they were also removed. As a consequence, leaf removal, regardless of timing, resulted in fewer laterals in the fruit zone (Figure 2). However, in 2013, prebloom leaf removal occurred prior to the initial (albeit small) development of lateral shoots and in both varieties, this treatment had more lateral shoots developing than the bloom treatment (i.e., fewer instances of a “no lateral shoot present” rating; Figure 2). To obtain a partial canopy refill but still maintain the benefits of FZLR, these results suggest that prebloom leaf removal is likely the optimal timing under the environmental conditions presented here.
One challenge to producing aromatic varieties such as Riesling and Sauvignon blanc in warm climates can be loss of varietal characteristics due to excessive heating of the fruit. Compounding the macroclimate effects are practices such as FZLR that expose clusters to sunlight and thus, higher fruit temperatures (Spayd et al. 2002). FZLR increases accumulation of these aromatic compounds (Vilanova et al. 2012, Zoecklein et al. 1998). In this study, bloom leaf removal in Riesling increased terpene concentrations (specifically, α-ionone and nerol oxide), likely due to the increased sun exposure (Zoecklein et al. 1998). Prebloom leaf removal in Riesling reduced aldehydes; past studies showed high concentrations of aldehydes in shaded fruit (Lohitnavy et al. 2010) and thus, the reduced aldehydes seen here may be due to increased exposure of the fruit. Aldehydes are often associated with an herbaceous or grassy aroma, while terpenes are associated with floral aromas (Rapp and Mandery 1986, Ristic et al. 2007, Simpson 1978). Our results indicate that the timing of leaf removal may influence the aromatic character of the fruit and thus, growers may tailor their timing to meet their needs and preferred wine styles: prebloom leaf removal improved floral character, while bloom leaf removal reduced grass character.
Washington grapes are characteristically lower in free ammonia than grapes from other regions (Spayd and Andersen-Bagge 1996). This can pose problems for yeast nutrition during fermentation. Typically, free ammonia concentrations from 150 to 400 mg/L are required for successful fermentation, but the number can vary based on grape variety and yeast strain used (Ugliano et al. 2007). The present study indicated that the timing of FZLR can impact total free ammonia concentration. While all treatments, control included, had the low free nitrogen concentration typically seen in eastern Washington, only the prebloom leaf removal treatment consistently did not result in lower free ammonia than the control. However, reduced total ammonia as a result of leaf removal may not be of concern to conventional growers, as they would still likely require ammonia additions in the winery for successful fermentation. The reduced free ammonia as a result of early leaf removal might be a consideration for organic wine production, where sources for nitrogen additions for yeast are more limited.
The desired tannin and phenolic concentrations differ among wine styles, but in general, higher concentrations are desirable in red wines and lower levels in white wines. Prebloom and bloom leaf removal increased skin tannin and phenolic concentrations in Sauvignon blanc, depending on the year. The increased skin tannins likely result from increased cluster exposure (Ristic et al. 2007). In 2012, the prebloom treatment had more tannins and phenolics than the control, while the bloom treatment had these same effects in 2013. Interestingly, in both years, the prebloom and bloom treatments had similar skin tannin and phenolic concentrations. However, it is unlikely that the increased skin tannin and phenolic concentrations as a result of leaf removal would translate through to the wine because standard white wine processing procedures allow little to no skin and seed contact.
Conclusion
This study suggests that leaf removal prior to and during bloom is an appropriate cultural practice for eastern Washington winegrape production of the region’s flagship white varieties, Sauvignon blanc and Riesling. FZLR during these times optimized both horticultural and disease management attributes of the practice without reducing fruit set or increasing sunburn. Additional enological properties, such as increased terpenes and reduced aldehydes, can be altered depending on the timing selected. Overall, this study suggests that the current practice of FZLR between fruit set and bunch closure may be improved by advancing the implementation window to earlier in the growing season. However, with the currently available vineyard technology, this cultural practice would most likely be implemented using hand labor, and the costs of this should be weighed against the potential improvements in spray coverage or juice aromatic characteristics.
Acknowledgments
Acknowledgments: This research was supported by the Washington State Grape and Wine Research Program and Mercer Estates in Prosser, WA. The authors thank Rick Hamman of Hogue Ranches for vineyard coordination and manuscript review and Maria Mireles, Scott Mattinson, Jensena Newhouse, and Steve Hoff of Washington State University for technical assistance. The authors also thank Gary Grove, Washington State University, for his review of the manuscript. The research described here represents a portion of a thesis submitted by B.L. Komm to the Graduate School of Washington State University in partial fulfillment of the requirements for an M.S. in Horticulture.
- Received January 2015.
- Revision received May 2015.
- Revision received July 2015.
- Accepted August 2015.
- Published online October 2015
This is an open access article distributed under the CC BY license (https://creativecommons.org/licenses/by/4.0/).
Literature Cited
Vol 66 Issue 4








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
More from this TOC section
Similar Articles
