





Abstract
The plant hormone abscisic acid (ABA) induces a developmental switch in the aquatic fern Marsilea quadrifolia, causing the formation of aerial type characteristics, including the elongation of petioles and roots, a change in leaf morphology, the expansion of leaf surface area, and the shortening of the internodes. A number of ABA-responsive heterophylly (ABRH) genes are induced early during the transition. Using optically pure isomers of ABA, it was found that both the natural S-(+)-ABA and the unnatural R-(−)-ABA are capable of inducing a heterophyllous switch and regulating ABRH gene expression. When dose responses are compared, the unnatural ABA gives stronger morphogenic effects than the natural ABA at the same concentration, it is effective at lower concentrations, and its optimal concentration is also lower compared with the natural ABA. Deuterium-labelled ABA enantiomers were used to trace the fate of applied ABA and to distinguish the applied compound and its metabolites from the endogenous counterparts. In tissues, the supplied (+)-ABA was metabolized principally to dihydrophaseic acid, while the supplied (−)-ABA was converted at a slower rate to 7′-hydroxy abscisic acid. Treatment with either enantiomer resulted in increased biosynthesis of ABA, as reflected in the accumulation of endogenous dihydrophaseic acid. Taken together, these results suggest two distinct mechanisms of action for (−)-ABA: either (−)-ABA is intrinsically active, or its activity is due to the stimulation of ABA biosynthesis.
This paper is available online free of all access charges (see http://jxb.oxfordjournals.org/open_access.html for further details)
The online version of this article has been published under an Open Access model. Users are entitled to use, reproduce, disseminate, or display the Open Access version of this article for non-commercial purposes provided that: the original authorship is properly and fully attributed; the Journal and the Society for Experimental Biology are attributed as the original place of publication with the correct citation details given; if an article is subsequently reproduced or disseminated not in its entirety but only in part or as a derivative work this must be clearly indicated. For commercial re-use, please contact: journals.permissions@oxfordjournals.org
Introduction
Heterophylly in aquatic plants serves an adaptive advantage, and has been a subject of investigation for fundamental mechanisms in plant morphogenesis (Gifford and Foster, 1988; Steeves and Sussex, 1989; Trewavas and Jones, 1991). Controlled by the developmental programme, environmental factors or both, plants produce distinct types of leaves corresponding to changes in water level and the seasons. In the laboratory, heterophylly is affected by cultural parameters. When conditions are otherwise in favour of producing the submerged type morphology, an exogenous supply of the plant hormone abscisic acid (ABA) at an optimal concentration induces aerial type characteristics (Zeevaart and Creelman, 1988; Lin and Yang, 1999). This effect is seen in a number of phylogenetically divergent aquatic plants. In Marsilea quadrifolia, heterophylly is also affected by gibberellins (Allsopp, 1962), phytochrome (Gaudet, 1963), and blue light (Gaudet, 1965; Lin and Yang, 1999).
This raises the question whether applied ABA mimics environmental factors or it is truly the endogenous signal for the heterophyllous switch (Trewavas and Jones, 1991). It has been shown previously that the blue light signal was not mediated by ABA, but that a change in culture medium composition was (Lin and Yang, 1999). Others demonstrated that osmotic stress and high photon fluence caused similar effects that could be correlated with ABA (Goliber, 1989; Goliber and Feldman, 1989). However, low cell turgor and ABA seemed to induce aerial morphology via different mechanisms (Deschamp and Cooke, 1983, 1984).
Heterophylly is the most morphologically distinct effect of ABA during the vegetative phase. In M. quadrifolia ABA has differential effects on organ development that are both qualitative and quantitative (Liu, 1984; Lin and Yang, 1999; Hsu et al., 2001). It dramatically promotes growth in leaves and roots, but inhibits growth in the internodes. The responses are progressive and dose-dependent. ABA has to be continuously present for the completion of the morphogenesis; when ABA is removed, growth and development return to the default mode (Hsu et al., 2001).
Growth analyses indicate that, for heterophyllous induction, there is a developmental window of responsiveness; only the primordial tissues are affected by ABA (Hsu et al., 2001). Since all organs of the mature M. quadrifolia plant are derived from the shoot apical meristem, the tissues responsive to ABA are localized in the shoot apex. From shoot apices, 24 ABA-responsive early genes, the ABRHs, were isolated; seven of them are immediate early genes (Hsu et al., 2001). The ABA-induced growth characteristics and the ABRHs provide useful developmental and molecular markers for the dissection of ABA-mediated signal transduction.
Heterophylly studies in M. quadrifolia have been performed to date using commercially available ABA. This ABA is a product of chemical synthesis and is racemic, i.e. consisting of a 1:1 mixture of the enatiomers, namely the natural hormone, S-(+)-ABA, and its mirror image, R-(−)-ABA, which has not been found in plant tissues. In numerous previous studies the unnatural enantiomer has been observed to elicit biological activity in some but not all ABA responsive processes. Table 1 summarizes the researchers' interpretation of the results of the biological activities that have been examined using optically pure enantiomers. It shows that the structural requirement of ABA action varies depending on the experimental system, the developmental stage, the cell type, the concentration used, and the time-course. For example, (+)- and (−)-ABA were found to be equally active in inhibiting germination of wheat embryos (Milborrow, 1970; Walker-Simmons et al., 1992; Rose et al., 1996), for inducing stomatal closure in barley leaves (Cummins et al., 1971), and for interfering with the synthesis and release of α-amylase induced by gibberellin in barley half seeds (Sondheimer et al., 1971). Unnatural (−)-ABA was found less effective in inhibiting the germination of barley seeds and excised ash embryos (Sondheimer et al., 1971), in increasing freezing tolerance in bromegrass suspension cell culture (Robertson et al., 1994), and in inhibiting the growth of maize suspension cells (Balsevich et al., 1994b). Such is also the case for the regulation of gene expression. For example, both (+)- and (−)-ABA were found to induce the expression of dhn (rab) and group 3 lea in imbibed wheat embryo, but the Em gene was only induced by the natural ABA (Walker-Simmons et al., 1992). In Arabidopsis thaliana cell cultures (+)-ABA and not (−)-ABA induced RAB18 gene expression (Jeannette et al., 1999).
Comparison of biological effects of exogenously applied (+)- and (−)-ABA
Effect | Biological activity | Target tissue/stage | Plant | Concentration used (μM) | Reference | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Equally effective | ||||||||||
| Inhibition | GA-induced synthesis and release of α-amylase | Embryo-free half seed | Barley | 0.1–1 | Sondheimer et al., 1971 | |||||
| GA-induced synthesis of α-amylase | Aleurone layers | Barley | 0–20 | Abrams et al., 1993; Hill et al., 1995 | ||||||
| Aleurone protoplasts | Hill et al., 1995 | |||||||||
| Synthesis and release of α-amylase | Embryo-containing half seed | Barley | 0.1–1 | Sondheimer et al., 1971 | ||||||
| Germination | Dormant seed embryo | Wheat | 1 and 10 | Rose et al., 1996; Walker-Simmons et al., 1992 | ||||||
| Germination | Isolated seed embryo | Barley | 1–5 | Abrams et al., 1993 | ||||||
| Growth | Coleoptile | Wheat | ? | Milborrow, 1970 | ||||||
| Growth | 8–16 fronds plantlets | Duckweed | 0.125 | Smart et al., 1995 | ||||||
| Growth | Microspore derived embryo | Oil seed rape | 1–10 | Wilmer et al., 1998 | ||||||
| Carrier-mediated uptake of (+)-ABA | Suspension culture cells | Carrot | 0–30 | Windsor et al., 1994 | ||||||
| Carrier-mediated uptake of ABA | Suspension culture cells | Amaranthus tricolor | 0–35 | Bianco-Colomas et al., 1991 | ||||||
| Promotion | Abscission assay | Explant | Cotton | 0.01–1.0 μg petiole−1 | Sondheimer et al., 1971 | |||||
| ABA 8′-hydroxylase induction | Cell suspension culture | Potato, Arabidopsis | 50 | Windsor and Zeevaart, 1997 | ||||||
| Desiccation tolerance, callus viability, LEA gene expression | Callus | Craterostigma plantagineum | 20 | Chandler et al., 1997 | ||||||
| Elongase activity | Microspore-derived embryo | Oilseed rape | 10 | Wilmer et al., 1998 | ||||||
| Erucic acid content | Microspore-derived embryo | Oilseed rape | 1–10 | Wilmer et al., 1998 | ||||||
| Gene expression- LMW hsps | Immature somatic embryo | White spruce | 15 | Dong and Dunstan, 1996 | ||||||
| Gene expression- non-Lea-like genes: PgEM5, PgEMB23 | Embryogenic suspension culture | White spruce | 15 (1–3 h) | Dong and Dunstan, 1997 | ||||||
| Gene expression- dhn(rab), lea (group 3) | Embryo germination | Wheat | 10 | Walker-Simmons et al., 1992 | ||||||
| Gene expression- dehydrin | Suspension cell | Bromegrass | 25 | Wilen et al., 1996 | ||||||
| Growth | Excised axis | Bean | 17 | Sondheimer et al., 1971 | ||||||
| Protein accumulation- RAB | Suspension cell | Bromegrass | 25 | Wilen et al., 1996 | ||||||
| Stomatal closure | Leaf | Barley | 0.1 | Sondheimer et al., 1971 | ||||||
| Turion induction | 8–16 fronds plantlets | Duckweed | 0.125 | Smart et al., 1995 | ||||||
| (−)-ABA less effective | ||||||||||
| Inhibition | Germination | Dormant embryo | White ash | ? | Sondheimer et al., 1971 | |||||
| Germination | Non-dormant embryo | White ash | ? | Sondheimer et al., 1971 | ||||||
| Germination | Seed | Arabidopsis wild type, chilled | 0.3–5 | Nambara et al., 2002 | ||||||
| Germination | Seed | Cress | 0.1–100 | Gusta et al., 1992 | ||||||
| Germination | Seed | Tomato, lettuce | 1, 2, 5 and 10 | Toorop et al., 1999 | ||||||
| Germination | Seed | Lettuce | 0.1–100 | Nakano et al., 1995; Shen et al., 1995; Todoroki et al., 1994, 1995a, b | ||||||
| Germination | Microspore-derived embryo | Brassica napus | 0.05–1 | Hays et al., 1996 | ||||||
| Growth | Suspension cell | Maize | 10 | Balsevich et al., 1994b | ||||||
| Growth | Seedling | Barley | 3.8–38 | Sondheimer et al., 1971 | ||||||
| Growth | Seedling | Radish | 0.1–100 | Shen et al., 1995 | ||||||
| GA3-induced synthesis and release of α-amylase | Embryo-free half seed | Barley | 0.1–100 | Nakano et al., 1995; Todoroki et al., 1994, 1995a, b | ||||||
| Elongation | Third leaf | Maize | 1–10 | Cramer et al., 1998 | ||||||
| Elongation | Second leaf | Rice seedlings | 0.1–100 | Nakano et al., 1995; Todoroki et al., 1994, 1995a, b | ||||||
| Stomatal opening | Epidermal strips | Spiderwort | 10−4–1 | Nakano et al., 1995; Todoroki et al., 1994, 1995a, b | ||||||
| Transpiration rate | Seedling | Wheat | 1–1000 | Rose et al., 1996 | ||||||
| Transport of (+)-ABA by uptake carrier | Suspension cell cultures | Barley | 20 | Perras et al., 1994 | ||||||
| Promotion | Stomatal closure | Seedling | Barley | 8 | Cummins and Sondheimer, 1973 | |||||
| Freezing tolerance | Suspension cell | Bromegrass | 75; 25 and 75 | Robertson et al., 1994; Wilen et al., 1996 | ||||||
| 7′ and 8′-hydroxylase induction | Cell suspension cultures | Black Mexican sweet corn | 200 | Cutler et al., 1997 | ||||||
| Gene expression- wheat Em::uidA | Somatic embryo | White spruce | 24 | Bommineni et al., 1998 | ||||||
| Gene expression- wheat Em::uidA | Protoplast | White spruce | >100 | Dong et al., 1994 | ||||||
| Gene expression- Lea-like genes: PgEMB12, PgEMB14, PgEMB15 | Embryogenic suspension culture | White spruce | 15 (3–48 h) | Dong and Dunstan, 1997 | ||||||
| 10 and 100 (24 h) | ||||||||||
| Gene expression- non-Lea-like genes: PgEM5, PgEMB23 | Embryogenic suspension culture | White spruce | 10 and 100 (24 h) | Dong and Dunstan, 1997 | ||||||
| Gene expression- napin | Microspore-derived embryos | B. napus | 1, 10 and 30 | Wilen et al., 1993 | ||||||
| Gene expression- napin, oleosin | Microspore-derived embryos | B. napus | 1–30 | Hays et al., 1996 | ||||||
| Gene expression- GUS activity | Aleurone protoplasts | Barley | 0–10 | Hill et al., 1995 | ||||||
| Gene expression- GUS activity | Fresh seeds, developing transgenic seeds | Tobacco | 0–1 | Jiang et al., 1996 | ||||||
| Transport by uptake carrier | Suspension cells | Carrot | 0–30 | Windsor et al., 1994 | ||||||
| (−)-ABA no effect | ||||||||||
| Inhibition | Germination | Excised embryo | Yellow cedar | 1 | Schmitz et al., 2002 | |||||
| Stomatal opening | Isolated leaf epidermal strips, intact leaves | Commelina comunis, Tropaeolum majus | 100 | Milborrow, 1980 | ||||||
| Reduction | Osmotic potential | Suspension cell | Bromegrass | 25 | Wilen et al., 1996 | |||||
| Promotion | Gene expression- Em | Dormant seed embryo germination | Wheat | 10 | Walker-Simmons et al., 1992 | |||||
| Gene expression- WCS120 gene | Suspension cell culture | Barley | 20 | Perras et al., 1994 | ||||||
| Gene expression- RAB18 gene | Suspension cell culture | Arabidopsis | 10 | Jeannette et al., 1999 | ||||||
| pH transition in medium | Suspension cell | Maize | 10 | Balsevich et al., 1994b | ||||||
| Sucrose uptake | Suspension cell | Bromegrass | 25 | Wilen et al., 1996 | ||||||
| Maltose accumulation | Suspension cell | Bromegrass | 25 | Wilen et al., 1996 | ||||||
| Radial water transport | Root | Maize | 0.1 | Sauter et al., 2002 | ||||||
| Maturation | Somatic embryo | White spruce | 15 | Dunstan et al., 1992 | ||||||
Effect | Biological activity | Target tissue/stage | Plant | Concentration used (μM) | Reference | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Equally effective | ||||||||||
| Inhibition | GA-induced synthesis and release of α-amylase | Embryo-free half seed | Barley | 0.1–1 | Sondheimer et al., 1971 | |||||
| GA-induced synthesis of α-amylase | Aleurone layers | Barley | 0–20 | Abrams et al., 1993; Hill et al., 1995 | ||||||
| Aleurone protoplasts | Hill et al., 1995 | |||||||||
| Synthesis and release of α-amylase | Embryo-containing half seed | Barley | 0.1–1 | Sondheimer et al., 1971 | ||||||
| Germination | Dormant seed embryo | Wheat | 1 and 10 | Rose et al., 1996; Walker-Simmons et al., 1992 | ||||||
| Germination | Isolated seed embryo | Barley | 1–5 | Abrams et al., 1993 | ||||||
| Growth | Coleoptile | Wheat | ? | Milborrow, 1970 | ||||||
| Growth | 8–16 fronds plantlets | Duckweed | 0.125 | Smart et al., 1995 | ||||||
| Growth | Microspore derived embryo | Oil seed rape | 1–10 | Wilmer et al., 1998 | ||||||
| Carrier-mediated uptake of (+)-ABA | Suspension culture cells | Carrot | 0–30 | Windsor et al., 1994 | ||||||
| Carrier-mediated uptake of ABA | Suspension culture cells | Amaranthus tricolor | 0–35 | Bianco-Colomas et al., 1991 | ||||||
| Promotion | Abscission assay | Explant | Cotton | 0.01–1.0 μg petiole−1 | Sondheimer et al., 1971 | |||||
| ABA 8′-hydroxylase induction | Cell suspension culture | Potato, Arabidopsis | 50 | Windsor and Zeevaart, 1997 | ||||||
| Desiccation tolerance, callus viability, LEA gene expression | Callus | Craterostigma plantagineum | 20 | Chandler et al., 1997 | ||||||
| Elongase activity | Microspore-derived embryo | Oilseed rape | 10 | Wilmer et al., 1998 | ||||||
| Erucic acid content | Microspore-derived embryo | Oilseed rape | 1–10 | Wilmer et al., 1998 | ||||||
| Gene expression- LMW hsps | Immature somatic embryo | White spruce | 15 | Dong and Dunstan, 1996 | ||||||
| Gene expression- non-Lea-like genes: PgEM5, PgEMB23 | Embryogenic suspension culture | White spruce | 15 (1–3 h) | Dong and Dunstan, 1997 | ||||||
| Gene expression- dhn(rab), lea (group 3) | Embryo germination | Wheat | 10 | Walker-Simmons et al., 1992 | ||||||
| Gene expression- dehydrin | Suspension cell | Bromegrass | 25 | Wilen et al., 1996 | ||||||
| Growth | Excised axis | Bean | 17 | Sondheimer et al., 1971 | ||||||
| Protein accumulation- RAB | Suspension cell | Bromegrass | 25 | Wilen et al., 1996 | ||||||
| Stomatal closure | Leaf | Barley | 0.1 | Sondheimer et al., 1971 | ||||||
| Turion induction | 8–16 fronds plantlets | Duckweed | 0.125 | Smart et al., 1995 | ||||||
| (−)-ABA less effective | ||||||||||
| Inhibition | Germination | Dormant embryo | White ash | ? | Sondheimer et al., 1971 | |||||
| Germination | Non-dormant embryo | White ash | ? | Sondheimer et al., 1971 | ||||||
| Germination | Seed | Arabidopsis wild type, chilled | 0.3–5 | Nambara et al., 2002 | ||||||
| Germination | Seed | Cress | 0.1–100 | Gusta et al., 1992 | ||||||
| Germination | Seed | Tomato, lettuce | 1, 2, 5 and 10 | Toorop et al., 1999 | ||||||
| Germination | Seed | Lettuce | 0.1–100 | Nakano et al., 1995; Shen et al., 1995; Todoroki et al., 1994, 1995a, b | ||||||
| Germination | Microspore-derived embryo | Brassica napus | 0.05–1 | Hays et al., 1996 | ||||||
| Growth | Suspension cell | Maize | 10 | Balsevich et al., 1994b | ||||||
| Growth | Seedling | Barley | 3.8–38 | Sondheimer et al., 1971 | ||||||
| Growth | Seedling | Radish | 0.1–100 | Shen et al., 1995 | ||||||
| GA3-induced synthesis and release of α-amylase | Embryo-free half seed | Barley | 0.1–100 | Nakano et al., 1995; Todoroki et al., 1994, 1995a, b | ||||||
| Elongation | Third leaf | Maize | 1–10 | Cramer et al., 1998 | ||||||
| Elongation | Second leaf | Rice seedlings | 0.1–100 | Nakano et al., 1995; Todoroki et al., 1994, 1995a, b | ||||||
| Stomatal opening | Epidermal strips | Spiderwort | 10−4–1 | Nakano et al., 1995; Todoroki et al., 1994, 1995a, b | ||||||
| Transpiration rate | Seedling | Wheat | 1–1000 | Rose et al., 1996 | ||||||
| Transport of (+)-ABA by uptake carrier | Suspension cell cultures | Barley | 20 | Perras et al., 1994 | ||||||
| Promotion | Stomatal closure | Seedling | Barley | 8 | Cummins and Sondheimer, 1973 | |||||
| Freezing tolerance | Suspension cell | Bromegrass | 75; 25 and 75 | Robertson et al., 1994; Wilen et al., 1996 | ||||||
| 7′ and 8′-hydroxylase induction | Cell suspension cultures | Black Mexican sweet corn | 200 | Cutler et al., 1997 | ||||||
| Gene expression- wheat Em::uidA | Somatic embryo | White spruce | 24 | Bommineni et al., 1998 | ||||||
| Gene expression- wheat Em::uidA | Protoplast | White spruce | >100 | Dong et al., 1994 | ||||||
| Gene expression- Lea-like genes: PgEMB12, PgEMB14, PgEMB15 | Embryogenic suspension culture | White spruce | 15 (3–48 h) | Dong and Dunstan, 1997 | ||||||
| 10 and 100 (24 h) | ||||||||||
| Gene expression- non-Lea-like genes: PgEM5, PgEMB23 | Embryogenic suspension culture | White spruce | 10 and 100 (24 h) | Dong and Dunstan, 1997 | ||||||
| Gene expression- napin | Microspore-derived embryos | B. napus | 1, 10 and 30 | Wilen et al., 1993 | ||||||
| Gene expression- napin, oleosin | Microspore-derived embryos | B. napus | 1–30 | Hays et al., 1996 | ||||||
| Gene expression- GUS activity | Aleurone protoplasts | Barley | 0–10 | Hill et al., 1995 | ||||||
| Gene expression- GUS activity | Fresh seeds, developing transgenic seeds | Tobacco | 0–1 | Jiang et al., 1996 | ||||||
| Transport by uptake carrier | Suspension cells | Carrot | 0–30 | Windsor et al., 1994 | ||||||
| (−)-ABA no effect | ||||||||||
| Inhibition | Germination | Excised embryo | Yellow cedar | 1 | Schmitz et al., 2002 | |||||
| Stomatal opening | Isolated leaf epidermal strips, intact leaves | Commelina comunis, Tropaeolum majus | 100 | Milborrow, 1980 | ||||||
| Reduction | Osmotic potential | Suspension cell | Bromegrass | 25 | Wilen et al., 1996 | |||||
| Promotion | Gene expression- Em | Dormant seed embryo germination | Wheat | 10 | Walker-Simmons et al., 1992 | |||||
| Gene expression- WCS120 gene | Suspension cell culture | Barley | 20 | Perras et al., 1994 | ||||||
| Gene expression- RAB18 gene | Suspension cell culture | Arabidopsis | 10 | Jeannette et al., 1999 | ||||||
| pH transition in medium | Suspension cell | Maize | 10 | Balsevich et al., 1994b | ||||||
| Sucrose uptake | Suspension cell | Bromegrass | 25 | Wilen et al., 1996 | ||||||
| Maltose accumulation | Suspension cell | Bromegrass | 25 | Wilen et al., 1996 | ||||||
| Radial water transport | Root | Maize | 0.1 | Sauter et al., 2002 | ||||||
| Maturation | Somatic embryo | White spruce | 15 | Dunstan et al., 1992 | ||||||
Comparison of biological effects of exogenously applied (+)- and (−)-ABA
Effect | Biological activity | Target tissue/stage | Plant | Concentration used (μM) | Reference | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Equally effective | ||||||||||
| Inhibition | GA-induced synthesis and release of α-amylase | Embryo-free half seed | Barley | 0.1–1 | Sondheimer et al., 1971 | |||||
| GA-induced synthesis of α-amylase | Aleurone layers | Barley | 0–20 | Abrams et al., 1993; Hill et al., 1995 | ||||||
| Aleurone protoplasts | Hill et al., 1995 | |||||||||
| Synthesis and release of α-amylase | Embryo-containing half seed | Barley | 0.1–1 | Sondheimer et al., 1971 | ||||||
| Germination | Dormant seed embryo | Wheat | 1 and 10 | Rose et al., 1996; Walker-Simmons et al., 1992 | ||||||
| Germination | Isolated seed embryo | Barley | 1–5 | Abrams et al., 1993 | ||||||
| Growth | Coleoptile | Wheat | ? | Milborrow, 1970 | ||||||
| Growth | 8–16 fronds plantlets | Duckweed | 0.125 | Smart et al., 1995 | ||||||
| Growth | Microspore derived embryo | Oil seed rape | 1–10 | Wilmer et al., 1998 | ||||||
| Carrier-mediated uptake of (+)-ABA | Suspension culture cells | Carrot | 0–30 | Windsor et al., 1994 | ||||||
| Carrier-mediated uptake of ABA | Suspension culture cells | Amaranthus tricolor | 0–35 | Bianco-Colomas et al., 1991 | ||||||
| Promotion | Abscission assay | Explant | Cotton | 0.01–1.0 μg petiole−1 | Sondheimer et al., 1971 | |||||
| ABA 8′-hydroxylase induction | Cell suspension culture | Potato, Arabidopsis | 50 | Windsor and Zeevaart, 1997 | ||||||
| Desiccation tolerance, callus viability, LEA gene expression | Callus | Craterostigma plantagineum | 20 | Chandler et al., 1997 | ||||||
| Elongase activity | Microspore-derived embryo | Oilseed rape | 10 | Wilmer et al., 1998 | ||||||
| Erucic acid content | Microspore-derived embryo | Oilseed rape | 1–10 | Wilmer et al., 1998 | ||||||
| Gene expression- LMW hsps | Immature somatic embryo | White spruce | 15 | Dong and Dunstan, 1996 | ||||||
| Gene expression- non-Lea-like genes: PgEM5, PgEMB23 | Embryogenic suspension culture | White spruce | 15 (1–3 h) | Dong and Dunstan, 1997 | ||||||
| Gene expression- dhn(rab), lea (group 3) | Embryo germination | Wheat | 10 | Walker-Simmons et al., 1992 | ||||||
| Gene expression- dehydrin | Suspension cell | Bromegrass | 25 | Wilen et al., 1996 | ||||||
| Growth | Excised axis | Bean | 17 | Sondheimer et al., 1971 | ||||||
| Protein accumulation- RAB | Suspension cell | Bromegrass | 25 | Wilen et al., 1996 | ||||||
| Stomatal closure | Leaf | Barley | 0.1 | Sondheimer et al., 1971 | ||||||
| Turion induction | 8–16 fronds plantlets | Duckweed | 0.125 | Smart et al., 1995 | ||||||
| (−)-ABA less effective | ||||||||||
| Inhibition | Germination | Dormant embryo | White ash | ? | Sondheimer et al., 1971 | |||||
| Germination | Non-dormant embryo | White ash | ? | Sondheimer et al., 1971 | ||||||
| Germination | Seed | Arabidopsis wild type, chilled | 0.3–5 | Nambara et al., 2002 | ||||||
| Germination | Seed | Cress | 0.1–100 | Gusta et al., 1992 | ||||||
| Germination | Seed | Tomato, lettuce | 1, 2, 5 and 10 | Toorop et al., 1999 | ||||||
| Germination | Seed | Lettuce | 0.1–100 | Nakano et al., 1995; Shen et al., 1995; Todoroki et al., 1994, 1995a, b | ||||||
| Germination | Microspore-derived embryo | Brassica napus | 0.05–1 | Hays et al., 1996 | ||||||
| Growth | Suspension cell | Maize | 10 | Balsevich et al., 1994b | ||||||
| Growth | Seedling | Barley | 3.8–38 | Sondheimer et al., 1971 | ||||||
| Growth | Seedling | Radish | 0.1–100 | Shen et al., 1995 | ||||||
| GA3-induced synthesis and release of α-amylase | Embryo-free half seed | Barley | 0.1–100 | Nakano et al., 1995; Todoroki et al., 1994, 1995a, b | ||||||
| Elongation | Third leaf | Maize | 1–10 | Cramer et al., 1998 | ||||||
| Elongation | Second leaf | Rice seedlings | 0.1–100 | Nakano et al., 1995; Todoroki et al., 1994, 1995a, b | ||||||
| Stomatal opening | Epidermal strips | Spiderwort | 10−4–1 | Nakano et al., 1995; Todoroki et al., 1994, 1995a, b | ||||||
| Transpiration rate | Seedling | Wheat | 1–1000 | Rose et al., 1996 | ||||||
| Transport of (+)-ABA by uptake carrier | Suspension cell cultures | Barley | 20 | Perras et al., 1994 | ||||||
| Promotion | Stomatal closure | Seedling | Barley | 8 | Cummins and Sondheimer, 1973 | |||||
| Freezing tolerance | Suspension cell | Bromegrass | 75; 25 and 75 | Robertson et al., 1994; Wilen et al., 1996 | ||||||
| 7′ and 8′-hydroxylase induction | Cell suspension cultures | Black Mexican sweet corn | 200 | Cutler et al., 1997 | ||||||
| Gene expression- wheat Em::uidA | Somatic embryo | White spruce | 24 | Bommineni et al., 1998 | ||||||
| Gene expression- wheat Em::uidA | Protoplast | White spruce | >100 | Dong et al., 1994 | ||||||
| Gene expression- Lea-like genes: PgEMB12, PgEMB14, PgEMB15 | Embryogenic suspension culture | White spruce | 15 (3–48 h) | Dong and Dunstan, 1997 | ||||||
| 10 and 100 (24 h) | ||||||||||
| Gene expression- non-Lea-like genes: PgEM5, PgEMB23 | Embryogenic suspension culture | White spruce | 10 and 100 (24 h) | Dong and Dunstan, 1997 | ||||||
| Gene expression- napin | Microspore-derived embryos | B. napus | 1, 10 and 30 | Wilen et al., 1993 | ||||||
| Gene expression- napin, oleosin | Microspore-derived embryos | B. napus | 1–30 | Hays et al., 1996 | ||||||
| Gene expression- GUS activity | Aleurone protoplasts | Barley | 0–10 | Hill et al., 1995 | ||||||
| Gene expression- GUS activity | Fresh seeds, developing transgenic seeds | Tobacco | 0–1 | Jiang et al., 1996 | ||||||
| Transport by uptake carrier | Suspension cells | Carrot | 0–30 | Windsor et al., 1994 | ||||||
| (−)-ABA no effect | ||||||||||
| Inhibition | Germination | Excised embryo | Yellow cedar | 1 | Schmitz et al., 2002 | |||||
| Stomatal opening | Isolated leaf epidermal strips, intact leaves | Commelina comunis, Tropaeolum majus | 100 | Milborrow, 1980 | ||||||
| Reduction | Osmotic potential | Suspension cell | Bromegrass | 25 | Wilen et al., 1996 | |||||
| Promotion | Gene expression- Em | Dormant seed embryo germination | Wheat | 10 | Walker-Simmons et al., 1992 | |||||
| Gene expression- WCS120 gene | Suspension cell culture | Barley | 20 | Perras et al., 1994 | ||||||
| Gene expression- RAB18 gene | Suspension cell culture | Arabidopsis | 10 | Jeannette et al., 1999 | ||||||
| pH transition in medium | Suspension cell | Maize | 10 | Balsevich et al., 1994b | ||||||
| Sucrose uptake | Suspension cell | Bromegrass | 25 | Wilen et al., 1996 | ||||||
| Maltose accumulation | Suspension cell | Bromegrass | 25 | Wilen et al., 1996 | ||||||
| Radial water transport | Root | Maize | 0.1 | Sauter et al., 2002 | ||||||
| Maturation | Somatic embryo | White spruce | 15 | Dunstan et al., 1992 | ||||||
Effect | Biological activity | Target tissue/stage | Plant | Concentration used (μM) | Reference | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Equally effective | ||||||||||
| Inhibition | GA-induced synthesis and release of α-amylase | Embryo-free half seed | Barley | 0.1–1 | Sondheimer et al., 1971 | |||||
| GA-induced synthesis of α-amylase | Aleurone layers | Barley | 0–20 | Abrams et al., 1993; Hill et al., 1995 | ||||||
| Aleurone protoplasts | Hill et al., 1995 | |||||||||
| Synthesis and release of α-amylase | Embryo-containing half seed | Barley | 0.1–1 | Sondheimer et al., 1971 | ||||||
| Germination | Dormant seed embryo | Wheat | 1 and 10 | Rose et al., 1996; Walker-Simmons et al., 1992 | ||||||
| Germination | Isolated seed embryo | Barley | 1–5 | Abrams et al., 1993 | ||||||
| Growth | Coleoptile | Wheat | ? | Milborrow, 1970 | ||||||
| Growth | 8–16 fronds plantlets | Duckweed | 0.125 | Smart et al., 1995 | ||||||
| Growth | Microspore derived embryo | Oil seed rape | 1–10 | Wilmer et al., 1998 | ||||||
| Carrier-mediated uptake of (+)-ABA | Suspension culture cells | Carrot | 0–30 | Windsor et al., 1994 | ||||||
| Carrier-mediated uptake of ABA | Suspension culture cells | Amaranthus tricolor | 0–35 | Bianco-Colomas et al., 1991 | ||||||
| Promotion | Abscission assay | Explant | Cotton | 0.01–1.0 μg petiole−1 | Sondheimer et al., 1971 | |||||
| ABA 8′-hydroxylase induction | Cell suspension culture | Potato, Arabidopsis | 50 | Windsor and Zeevaart, 1997 | ||||||
| Desiccation tolerance, callus viability, LEA gene expression | Callus | Craterostigma plantagineum | 20 | Chandler et al., 1997 | ||||||
| Elongase activity | Microspore-derived embryo | Oilseed rape | 10 | Wilmer et al., 1998 | ||||||
| Erucic acid content | Microspore-derived embryo | Oilseed rape | 1–10 | Wilmer et al., 1998 | ||||||
| Gene expression- LMW hsps | Immature somatic embryo | White spruce | 15 | Dong and Dunstan, 1996 | ||||||
| Gene expression- non-Lea-like genes: PgEM5, PgEMB23 | Embryogenic suspension culture | White spruce | 15 (1–3 h) | Dong and Dunstan, 1997 | ||||||
| Gene expression- dhn(rab), lea (group 3) | Embryo germination | Wheat | 10 | Walker-Simmons et al., 1992 | ||||||
| Gene expression- dehydrin | Suspension cell | Bromegrass | 25 | Wilen et al., 1996 | ||||||
| Growth | Excised axis | Bean | 17 | Sondheimer et al., 1971 | ||||||
| Protein accumulation- RAB | Suspension cell | Bromegrass | 25 | Wilen et al., 1996 | ||||||
| Stomatal closure | Leaf | Barley | 0.1 | Sondheimer et al., 1971 | ||||||
| Turion induction | 8–16 fronds plantlets | Duckweed | 0.125 | Smart et al., 1995 | ||||||
| (−)-ABA less effective | ||||||||||
| Inhibition | Germination | Dormant embryo | White ash | ? | Sondheimer et al., 1971 | |||||
| Germination | Non-dormant embryo | White ash | ? | Sondheimer et al., 1971 | ||||||
| Germination | Seed | Arabidopsis wild type, chilled | 0.3–5 | Nambara et al., 2002 | ||||||
| Germination | Seed | Cress | 0.1–100 | Gusta et al., 1992 | ||||||
| Germination | Seed | Tomato, lettuce | 1, 2, 5 and 10 | Toorop et al., 1999 | ||||||
| Germination | Seed | Lettuce | 0.1–100 | Nakano et al., 1995; Shen et al., 1995; Todoroki et al., 1994, 1995a, b | ||||||
| Germination | Microspore-derived embryo | Brassica napus | 0.05–1 | Hays et al., 1996 | ||||||
| Growth | Suspension cell | Maize | 10 | Balsevich et al., 1994b | ||||||
| Growth | Seedling | Barley | 3.8–38 | Sondheimer et al., 1971 | ||||||
| Growth | Seedling | Radish | 0.1–100 | Shen et al., 1995 | ||||||
| GA3-induced synthesis and release of α-amylase | Embryo-free half seed | Barley | 0.1–100 | Nakano et al., 1995; Todoroki et al., 1994, 1995a, b | ||||||
| Elongation | Third leaf | Maize | 1–10 | Cramer et al., 1998 | ||||||
| Elongation | Second leaf | Rice seedlings | 0.1–100 | Nakano et al., 1995; Todoroki et al., 1994, 1995a, b | ||||||
| Stomatal opening | Epidermal strips | Spiderwort | 10−4–1 | Nakano et al., 1995; Todoroki et al., 1994, 1995a, b | ||||||
| Transpiration rate | Seedling | Wheat | 1–1000 | Rose et al., 1996 | ||||||
| Transport of (+)-ABA by uptake carrier | Suspension cell cultures | Barley | 20 | Perras et al., 1994 | ||||||
| Promotion | Stomatal closure | Seedling | Barley | 8 | Cummins and Sondheimer, 1973 | |||||
| Freezing tolerance | Suspension cell | Bromegrass | 75; 25 and 75 | Robertson et al., 1994; Wilen et al., 1996 | ||||||
| 7′ and 8′-hydroxylase induction | Cell suspension cultures | Black Mexican sweet corn | 200 | Cutler et al., 1997 | ||||||
| Gene expression- wheat Em::uidA | Somatic embryo | White spruce | 24 | Bommineni et al., 1998 | ||||||
| Gene expression- wheat Em::uidA | Protoplast | White spruce | >100 | Dong et al., 1994 | ||||||
| Gene expression- Lea-like genes: PgEMB12, PgEMB14, PgEMB15 | Embryogenic suspension culture | White spruce | 15 (3–48 h) | Dong and Dunstan, 1997 | ||||||
| 10 and 100 (24 h) | ||||||||||
| Gene expression- non-Lea-like genes: PgEM5, PgEMB23 | Embryogenic suspension culture | White spruce | 10 and 100 (24 h) | Dong and Dunstan, 1997 | ||||||
| Gene expression- napin | Microspore-derived embryos | B. napus | 1, 10 and 30 | Wilen et al., 1993 | ||||||
| Gene expression- napin, oleosin | Microspore-derived embryos | B. napus | 1–30 | Hays et al., 1996 | ||||||
| Gene expression- GUS activity | Aleurone protoplasts | Barley | 0–10 | Hill et al., 1995 | ||||||
| Gene expression- GUS activity | Fresh seeds, developing transgenic seeds | Tobacco | 0–1 | Jiang et al., 1996 | ||||||
| Transport by uptake carrier | Suspension cells | Carrot | 0–30 | Windsor et al., 1994 | ||||||
| (−)-ABA no effect | ||||||||||
| Inhibition | Germination | Excised embryo | Yellow cedar | 1 | Schmitz et al., 2002 | |||||
| Stomatal opening | Isolated leaf epidermal strips, intact leaves | Commelina comunis, Tropaeolum majus | 100 | Milborrow, 1980 | ||||||
| Reduction | Osmotic potential | Suspension cell | Bromegrass | 25 | Wilen et al., 1996 | |||||
| Promotion | Gene expression- Em | Dormant seed embryo germination | Wheat | 10 | Walker-Simmons et al., 1992 | |||||
| Gene expression- WCS120 gene | Suspension cell culture | Barley | 20 | Perras et al., 1994 | ||||||
| Gene expression- RAB18 gene | Suspension cell culture | Arabidopsis | 10 | Jeannette et al., 1999 | ||||||
| pH transition in medium | Suspension cell | Maize | 10 | Balsevich et al., 1994b | ||||||
| Sucrose uptake | Suspension cell | Bromegrass | 25 | Wilen et al., 1996 | ||||||
| Maltose accumulation | Suspension cell | Bromegrass | 25 | Wilen et al., 1996 | ||||||
| Radial water transport | Root | Maize | 0.1 | Sauter et al., 2002 | ||||||
| Maturation | Somatic embryo | White spruce | 15 | Dunstan et al., 1992 | ||||||
In a kinetic study of maize leaf elongation, Cramer et al. (1998) found that the ABA effect on growth inhibition was reversible. The leaf elongation rate was highly correlated with the steady-state internal ABA concentration of the growing zone. When compared on the basis of external concentration, the (−)-enantiomer of ABA had much less effect than (+)-ABA due to its low level of accumulation. From this analysis, these authors concluded that the leaf elongation rate was equally sensitive to internal concentrations of (+)- or (−)-ABA.
The concentration of ABA in plant tissues is the result of a combination of transport, biosynthetic, and catabolic processes (Cutler and Krochko, 1999). As shown in Fig. 1, the metabolism of natural ABA (1) occurs principally through oxidation of the 8′-methyl group, resulting in 8′-hydroxy ABA (2), which rearranges to phaseic acid (3, PA). Further reduction of PA leads to dihydrophaseic acid (4, DPA). Other metabolic processes include conjugation of the acids as glucose esters, as well as the hydroxylation of 7′-methyl (Hampson et al., 1992) and the 9′-methyl group of the ABA ring (Zhou et al., 2004). In plants, unnatural ABA (5) is oxidized to (−)-7′-hydroxy abscisic acid (6, 7′-OH ABA) as the major metabolic product (Hampson et al., 1992). While the biological activity of ABA is well documented, the effect of its metabolites has been less studied, due to the scarcity of the compounds. However, there are indications that the initial catabolites of ABA can have pronounced hormonal activity (Hill et al., 1995; Zou et al., 1995).
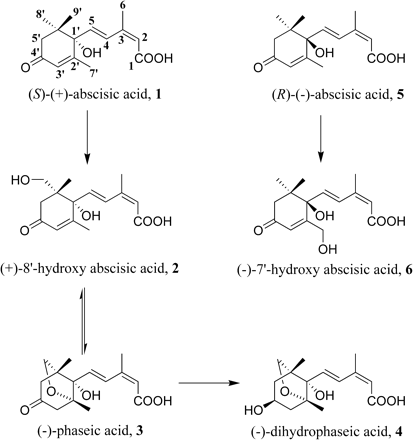
ABA and the main hydroxylated ABA metabolism products.
These results prompted an examination of the effects of optically pure isomers of ABA on heterophylly in M. quadrifolia. The main objective was to unravel the critical issue of structural requirements in ABA signal transduction, to relate effects of pure enantiomers to gene expression in order to determine which genes are involved in heterophylly. Previous studies showed that the continuous presence of ABA in the culture medium was critical for the maintenance of the heterophyllous switch (Hsu et al., 2001), and so determining the ABA concentrations in the tissues and the fate of supplied enantiomers may provide clues to the underlying mechanism. By feeding deuterium-labelled ABA enantiomers (Abrams et al., 2003) and tracing their fate in plant tissues, it was possible to distinguish the applied ABA from the endogenous hormone and to measure the changes in endogenous concentrations of the hormone and the metabolites. This is the first paper that reports the quantification of both supplied and endogenous ABA and its metabolites in plant tissues that could be correlated with a developmental switch induced by exogenous ABA.
Materials and methods
ABA enantiomers
The work related to the chemical preparation of ABA and ABA metabolites was carried out at the Plant Biotechnology Institute, National Research Council, Saskatoon, Canada. Thus, S-(+)-ABA and R-(−)-ABA were chemically synthesized and resolved into optically pure form by preparative HPLC resolution of racemic methyl abscisate followed by hydrolysis of the resolved esters as previously described (Dunstan et al., 1992). The (+)- and (−)-[4,5-d2]-ABA used in the feeding experiment, as well as the labelled internal standards used in liquid chromatography-electrospray ionization-tandem mass spectroscopy (LC-ES-MS-MS) analysis were either obtained by chemical synthesis (Abrams et al., 2003) or isolated as biotransformation products of ABA from maize cell suspension cultures (Zaharia et al., 2005).
Plant material and treatment
Aseptic cultures of Marsilea quadrifolia L. were established from sporocarps and propagated by subculturing into two-node segments (Liu, 1984; Lin and Yang, 1999). All plants used in this study were clones of the same plant. The basal medium is a liquid mineral medium (Laetsch and Briggs, 1961) supplemented with 3% sucrose. In all treatments and the control, the pH in the growth medium was adjusted to 6.4. ABA was added from a 1000× stock to the culture medium to reach the indicated final concentration. The treatment of ABA was applied 1 week after subculture. For the study of removal kinetics, plants were treated with ABA for 2 weeks, then the culture medium was removed and replaced with fresh basal medium without ABA. Plants were then allowed to grow for another 10 d. All cultures were kept in a growth room at 25 °C with a 16 h photoperiod. Illumination was provided with fluorescent tubes emitting near sunlight spectrum (FL40D-EX, Mitsubishi, Tokyo, Japan) at 40 μmol m−2 s−1.
RNA extraction and comparative RT-PCR
Total RNA was extracted from shoot apices using urea extraction buffer (Chen and Dellaporta, 1994), and precipitated in 2 M LiCl (Jepson et al., 1991). To be sure there is no contaminated DNA, the RNA preparation was further treated with DNase I. First strand cDNAs were synthesized with oligo(dT15) priming, serially diluted in water by a factor of ten, and then used as templates for PCR. The gene-specific primers previously identified (Hsu et al., 2001) were used for the detection of gene expression. PCR was carried out according to Sambrook et al. (1989), using 95 °C for 30 s, the Tm of gene-specific primers as the annealing temperature for 45 s, and 72 °C for 45 s, repeated for 35 cycles, then followed by 72 °C for 5 min. The RT-PCR products were resolved in 4% agarose gels (3% NuSieve and 1% SeaKem, FMC, Newport, Maine, USA), and the ethidium bromide-stained gel patterns were analysed in the Alpha Imager 1200 Documentation and Analysis System (Alpha Innotech, San Leandro, California, USA). For comparing the level of specific gene transcripts in shoot apices from plants in various treatments, the RT-PCR and gel analysis were carried out in batches, with the PCR amplification performed in serially diluted first strand cDNAs. A master reagent mix was prepared for each step of these analyses. Only those batches of RT-PCR showing consistent results in serially diluted templates were considered valid for determining the regulation pattern of gene expression. The comparative RT-PCR results were confirmed in at least two separate runs with each RNA sample and using at least two independent RNA sources.
Analysis of endogenous and applied ABA and ABA metabolites
This work was carried out at the Plant Biotechnology Institute, National Research Council, Saskatoon, Canada. Samples were taken each week during the 3-week incubation period. In the treatments and the control, three replicates were sampled, each with shoot apices pooled from eight plants. Plants were rinsed with sterile deionized water. After gently removing the excess water, the apex was excised from each plant and weighed. Samples were homogenized with a fine glass rod in liquid N2. To each sample a 40 μl mixture of water:acetonitrile (1:1, v/v) containing d4-ABA, d3-PA, d4-7′-OH ABA and d3-DPA (each at a concentration of 0.25 ng μl−1) was added. Further, a 1 ml mixture of isopropanol:glacial acetic acid (99:1, v/v) and a small magnetic stir bar were added to each sample. After 12 h of stirring in the dark, at room temperature, the samples were centrifuged at 1500 g for 10 min, the supernatant was transferred to a vial, and a second portion of 1 ml extraction solvent mixture was added. After stirring in the dark for another 4 h, the samples were centrifuged and the supernatant was combined with the previous one. The organic extract was dried under reduced pressure, then redissolved in 100 μl methanol:glacial acetic acid (99:1, v/v) followed by 900 ml of aqueous 1% glacial acetic acid. This mixture was further cleaned on the OASIS cartridge (Waters OASIS extraction cartridge HLB 1 cc). The fraction containing ABA and ABA metabolites was eluted with 1.5 ml methanol:water:glacial acetic acid (80:19:1, by vol.) and then evaporated to dryness. The final residue was dissolved in 200 μl (2×100 μl portions) mixture of water:acetonitrile (80:20, v/v) with 0.07% glacial acetic acid, and then subjected to LC-ES-MS-MS analysis, according to Ross et al. (2004) and Feurtado et al. (2004). For statistical analysis, the significance of the differences between sample means was calculated using the paired t-test.
Results
Both the natural (+)-ABA and the unnatural (−)-ABA induce heterophyllous switch and regulate ABRH gene expression
In the authors' laboratories, the cultural conditions were adjusted to sustain the formation of the submerged type of morphology (Fig. 2A). For the induction of heterophyllous transition, the optimal concentration of the commercial racemic ABA was 1 μM (Liu, 1984; Lin and Yang, 1999). Initially, the optically pure ABA was tested at this concentration, and it was found that both natural (+)-ABA and unnatural (−)-ABA are capable of inducing the developmental switch (compare Fig. 2A, B, and C). Typical aerial phenotypes are produced, the petioles and the roots elongate, the internodes shorten, the new leaves have expanded surface areas, and their morphology resembles four-leaf clover. Similar to the racemic ABA, the (+)- or (−)-ABA does not affect that part of the plant already developed prior to the treatment; the effects are only seen in the newly emerged organs, and are progressively stronger during the development. However, following the initial growth stimulation in the leaves and roots, prolonged ABA treatment triggers senescence, leading to developmental arrest and the transition to the reproductive phase. This is evident in the yellowing of the tissues, production of fewer nodes, and sporocarp formation in the ABA treated plants (Fig. 2B, C).
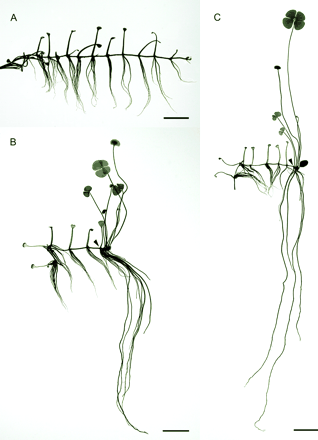
The effects of the optically pure ABA enantiomers on heterophylly in M. quadrifolia. (A) An untreated plant. (B) A plant treated with 1 μM S-(+)-ABA. (C) A plant treated with 1 μM R-(−)-ABA. In (B) and (C), the plants were grown in basal medium for 10 d, then treated with ABA for 4 weeks. Note that both ABA enantiomers induced heterophyllous switch. The R-(−)-ABA (C) had stronger effects than the S-(+)-ABA (B), resulting in longer petioles and roots, highly clustered nodes, as well as earlier leaf senescence and growth arrest. Arrowheads indicate the position of the shoot apex when ABA was added to the culture medium. Scale bars=1 cm.
In accord with the morphogenic responses, both ABA enantiomers also regulate the early responsive ABRH gene expression (Fig. 3). A number of ABRHs were examined, including the immediate early genes, ABRH1, ABRH3, ABRH4, ABRH5, and ABRH6, and the early secondary genes, ABRH10, ABRH12, ABRH18, ABRH19, and ABRH24. These genes are expressed at different levels and are regulated to different extents. Both enantiomers show comparable levels of regulation in most genes (Fig. 3). The encoded proteins include homologues of transcription factors, ABRH4, ABRH5, and ABRH12, homologues of signalling molecules, ABRH10 and ABRH18, a homologue of a membrane protein, ABRH6, and the chloroplast encoded proteins, ABRH19 and ABRH24 (Hsu et al., 2001). The regulation of an array of genes and the variety of gene identity, including regulatory and metabolic genes, suggest that both ABA enantiomers are associated with triggering the heterophyllous response in the morphogenic programme.
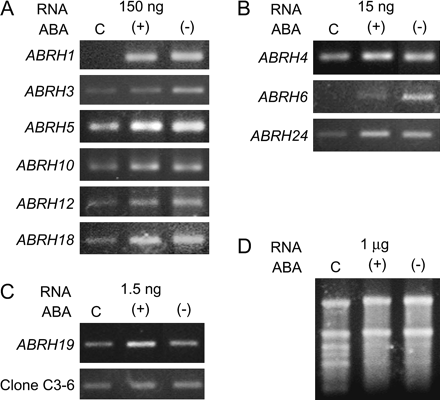
Both the natural (+)-ABA and the unnatural (−)-ABA regulate the early responsive ABRH gene expression in the shoot apices of M. quadrifolia. Plants were treated for 1 h with 1 μM ABA. ABA regulation is confirmed by a consistent pattern in comparative RT-PCR analyses using serially diluted RNA templates. Shown are representative gel images of RT-PCR products. Clone C3-6 is not regulated by ABA and serves as a control (Hsu et al., 2001). The RNA templates used are 150 ng, 15 ng, and 1.5 ng, respectively in (A), (B), and (C). The total RNA templates are shown in a gel image in (D). C, control, (+)=(+)-ABA-treated, (−)=(−)-ABA-treated.
The unnatural (−)-ABA has stronger effects than the natural (+)-ABA
Not only could both enantiomers induce the heterophyllous switch, but it was also observed that, initially, the growth responses of the plants were stronger when treated with (−)-ABA as compared to (+)-ABA (compare Fig. 2B and C), although later the differences appeared to be less pronounced. Therefore a series of growth kinetics studies was carried out to examine the time-course, the effect of ABA removal, and the dose–response, using as references length measurements of the various organs, including petioles, internodes, and roots.
In these studies, all the plants used were clones of the same plant, propagated from axillary buds of two-node segments (Liu, 1984; Hsu et al., 2001). At the time of subculture, the axillary buds often already contained the organ primordia for one to two nodes that, in response to ABA, develop into intermediate morphology. To minimize the residual effect on morphogenesis carried over in these primordia, plants were routinely used that developed three nodes, i.e. in these growth conditions 1 week after subculture (see the parts to the left of the arrowhead in Fig. 2B and C, and the nodes designated minus numbers in Figs 4–6).
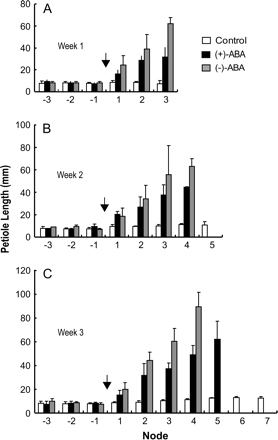
Time-course analyses of the effects of ABA enantiomers on petiole elongation. (A) Measurements after 1 week of treatment. (B) Measurements after 2 weeks of treatment. (C) Measurements after 3 weeks treatment. The concentration used was 1 μM. Arrows indicate the position of the shoot apex at the time of ABA application, and the leaf positions are labelled with reference to the treatment, i.e. 1 being the first leaf produced after the treatment, 2 being the second, etc. Note a progressive increase in petiole length during ABA treatment, and (−)-ABA has a stronger effect than (+)-ABA at each node. Immature leaves in young nodes are not shown, therefore a smaller number of leaves in the ABA-treated plants indicates an effect on growth arrest.
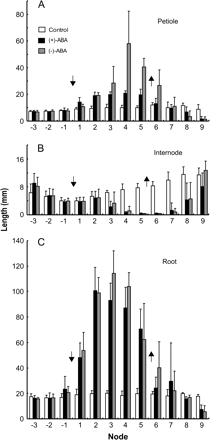
Growth kinetic responses to ABA application and removal. (A) The effects on petiole growth. (B) The effects on internode growth. (C) The effects on root growth. Plants were grown in basal medium for 1 week, treated with 1 μM of either ABA enantiomer for 2 weeks, then allowed to grow in fresh basal medium for another 10 d. The organs were measured at the end of the experimental period. A downward arrow indicates the position of the shoot apex at ABA application, and an upward arrow indicates that at ABA removal. ABA application induces aerial type characteristics, i.e. longer petioles and roots, and shorter internodes, whereas ABA removal reverses the effect. The effects are progressive upon ABA addition and removal.
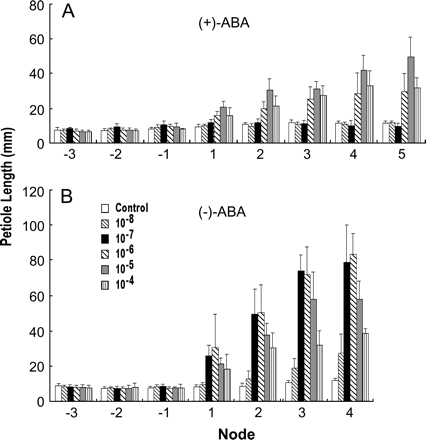
Dose-response of petiole elongation to exogenously supplied ABA enantiomers. (A) The responses to (+)-ABA. (B) The responses to (−)-ABA. Plants were grown in basal medium for 1 week, then treated with ABA at the indicated concentration for 3 weeks. Immature leaves are not shown.
Time-course of growth responses
Following ABA application, the plants produce new nodes bearing organs with striking changes in lengths compared with those existing before the treatment and in untreated plants (Figs 2, 4). Figure 4 gives the measurements of petiole growth. The effects are seen with both (+)-ABA and (−)-ABA, but the growth rates are higher in (−)-ABA. Table 2 compares the linear regression fit data for petiole growth curves, showing at least 2-fold difference in growth rate between treatments with (+)- and (−)-ABA at all time points.
Comparison of growth rates of petioles in ABA-treated plantsa
Treatment | Control | (+)-ABA | (−)-ABA | |
|---|---|---|---|---|
| Time-courseb | ||||
| Week 1 | 0.26±0.27 | 4.95±1.84 | 9.40±4.18 | |
| Week 2 | 0.53±0.15 | 5.01±1.89 | 13.63±1.34 | |
| Week 3 | 0.60±0.16 | 5.56±2.58 | 16.03±2.26 | |
| Reversibility kineticsc | ||||
| Application | 0.51±0.03 | 3.28±0.20 | 13.90±1.22 | |
| Removal | 0.30±0.09 | −3.65±0.33 | −11.40±0.64 |
Treatment | Control | (+)-ABA | (−)-ABA | |
|---|---|---|---|---|
| Time-courseb | ||||
| Week 1 | 0.26±0.27 | 4.95±1.84 | 9.40±4.18 | |
| Week 2 | 0.53±0.15 | 5.01±1.89 | 13.63±1.34 | |
| Week 3 | 0.60±0.16 | 5.56±2.58 | 16.03±2.26 | |
| Reversibility kineticsc | ||||
| Application | 0.51±0.03 | 3.28±0.20 | 13.90±1.22 | |
| Removal | 0.30±0.09 | −3.65±0.33 | −11.40±0.64 |
The slopes of the growth curves are shown as mean ±SE. The time interval for node development and leaf emergence is similar throughout the course of development in the treatments and the control. The growth rate was calculated using node development as a time unit.
The lengths of the petioles were measured at the end of each week. Paired t-test results for the difference in growth rates between the groups treated with (+)- and (−)-ABA are P <0.1 for week 1, P <0.02 for week 2, and P <0.001 for week 3 (n=3).
Plants were treated with 1 μM ABA for 2 weeks, then in fresh basal medium without ABA for 10 d. Petiole length was measured at the end of the experimental period. Paired t-test results for the difference in growth rates between (+)- and (−)-ABA treated groups are P <0.005 for ABA application, and P <0.007 for ABA removal (n=3).
Comparison of growth rates of petioles in ABA-treated plantsa
Treatment | Control | (+)-ABA | (−)-ABA | |
|---|---|---|---|---|
| Time-courseb | ||||
| Week 1 | 0.26±0.27 | 4.95±1.84 | 9.40±4.18 | |
| Week 2 | 0.53±0.15 | 5.01±1.89 | 13.63±1.34 | |
| Week 3 | 0.60±0.16 | 5.56±2.58 | 16.03±2.26 | |
| Reversibility kineticsc | ||||
| Application | 0.51±0.03 | 3.28±0.20 | 13.90±1.22 | |
| Removal | 0.30±0.09 | −3.65±0.33 | −11.40±0.64 |
Treatment | Control | (+)-ABA | (−)-ABA | |
|---|---|---|---|---|
| Time-courseb | ||||
| Week 1 | 0.26±0.27 | 4.95±1.84 | 9.40±4.18 | |
| Week 2 | 0.53±0.15 | 5.01±1.89 | 13.63±1.34 | |
| Week 3 | 0.60±0.16 | 5.56±2.58 | 16.03±2.26 | |
| Reversibility kineticsc | ||||
| Application | 0.51±0.03 | 3.28±0.20 | 13.90±1.22 | |
| Removal | 0.30±0.09 | −3.65±0.33 | −11.40±0.64 |
The slopes of the growth curves are shown as mean ±SE. The time interval for node development and leaf emergence is similar throughout the course of development in the treatments and the control. The growth rate was calculated using node development as a time unit.
The lengths of the petioles were measured at the end of each week. Paired t-test results for the difference in growth rates between the groups treated with (+)- and (−)-ABA are P <0.1 for week 1, P <0.02 for week 2, and P <0.001 for week 3 (n=3).
Plants were treated with 1 μM ABA for 2 weeks, then in fresh basal medium without ABA for 10 d. Petiole length was measured at the end of the experimental period. Paired t-test results for the difference in growth rates between (+)- and (−)-ABA treated groups are P <0.005 for ABA application, and P <0.007 for ABA removal (n=3).
The time-course study also shows that, in (−)-ABA treatments, the growth arrest has an early onset, such that fewer leaves are produced, compared with the untreated plants and those treated with (+)-ABA. While the growth persists in (+)-ABA treatments, some of the leaves produced later in (+)-ABA reach the length of the earlier leaves produced in (−)-ABA (Fig. 4). Hence it appears that the difference in the growth effects of the two ABA enantiomers becomes less distinct with prolonged treatment. The effects on root elongation and internode shortening are similarly progressive, as shown below.
The time-course of ABA effects was observed in more than a dozen experiments, either with the (+)- and (−)-ABA alone, or for a side-by-side comparison with the effects of their metabolites (H-J Wang, B-L Lin, unpublished results). Although the absolute values of organ measurements varied within a range, the progressively increasing or decreasing patterns of ABA responses were seen repeatedly.
Reversibility kinetics
It has previously been found that ABA has to be present continuously in order to complete the morphogenesis for heterophyllous switch (Hsu et al., 2001). When ABA is removed, intermediate morphology appears in organs that were already formed, yet still developing during the transition. To understand the action of optically pure ABA isomers, the growth kinetics responding to ABA removal were studied. Figure 5 shows that the progressive effect of ABA on organ development not only occurs during ABA treatment, but follows ABA removal as well. Moreover, the unnatural ABA has a distinctly stronger effect than the natural ABA on petiole growth (Fig. 5A). The rates of increase and decrease following ABA addition and removal, respectively, are 3–4-fold higher in the treatments with (−)-ABA than in those with (+)-ABA (Table 2).
The growth effects on internodes and roots are less distinguishable between the ABA enantiomers (Fig. 5B, C). The nodes are essentially clustered at the shoot apex, and the internode lengths are calculated from the total length of the rhizome fragment. Root growth extends for a long period of time and has been observed to continue for a month (Y-C Chang, R-W Chen, B-L Lin, unpublished results). During the experimental period in this study, 2 weeks for ABA treatment and 10 d after ABA removal, most roots did not complete growth to reach the maximal length.
Dose responses
The progressive and differential responses of the organs suggest a difference in the sensitivity of various developmental stages and of the tissues. This, together with the different intensity in the responses between ABA enantiomers, prompted an examination of the dosage effects. Concentrations spanning five orders of magnitude, from 10−8 M to 10−4 M, were used. Organ measurements were taken at the end of a 4-week treatment. Figure 6 shows that (+)-ABA was effective in promoting petiole elongation from 10−6 M to 10−4 M, with 10−5 M being the optimal concentration. Whereas (−)-ABA was effective from 10−8 M to 10−4 M, and the optimal concentration was between 10−7 M and 10−6 M.
Table 3 summarizes the dose responses. In each organ, the effective concentration and the optimal concentration of (+)- and (−)-ABA differ by one to two orders of magnitude. The unnatural ABA is effective at lower concentrations, the effective range is wider, and the optimal concentration is lower, compared with natural ABA. A low dosage effect was also noticed, i.e. the opposite response is produced at a concentration between the ineffective concentration and the lower end of the effective range. Among the organs, the root responds at lower concentrations than the leaf and the rhizome. In addition, the optimal concentration in the root shifts to a different level during the course of development, indicating biphasic responses.
Comparison of the effective concentrations of (+)-ABA and (−)-ABA on the induction of heterophyllous switch in M. quadrifolia
Organ | ABA isomer | Concentration (M) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Ineffective | Low dosage effecta | Effective range | Optimal | |||||||
| Petiole | (+) | 10−8 | 10−7 | 10−6∼10−4 | 10−5 | |||||
| (−) | – | – | 10−8∼10−4 | 10−7∼10−6b | ||||||
| Internode | (+) | 10−8 | 10−7 | 10−6∼10−4 | 10−5∼10−4b | |||||
| (−) | – | 10−8 | 10−7∼10−4 | ≥10−7c | ||||||
| Root | (+) | 10−8 | – | 10−7∼10−4 | Earlyd | Lated | ||||
| 10−5 | 10−6 | |||||||||
| (−) | – | – | 10−8∼10−4 | Earlyd | Lated | |||||
| 10−7∼10−6b | 10−5∼10−4b | |||||||||
Organ | ABA isomer | Concentration (M) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Ineffective | Low dosage effecta | Effective range | Optimal | |||||||
| Petiole | (+) | 10−8 | 10−7 | 10−6∼10−4 | 10−5 | |||||
| (−) | – | – | 10−8∼10−4 | 10−7∼10−6b | ||||||
| Internode | (+) | 10−8 | 10−7 | 10−6∼10−4 | 10−5∼10−4b | |||||
| (−) | – | 10−8 | 10−7∼10−4 | ≥10−7c | ||||||
| Root | (+) | 10−8 | – | 10−7∼10−4 | Earlyd | Lated | ||||
| 10−5 | 10−6 | |||||||||
| (−) | – | – | 10−8∼10−4 | Earlyd | Lated | |||||
| 10−7∼10−6b | 10−5∼10−4b | |||||||||
Plants were treated with the optically pure ABA enantiomer at various concentrations, ranging from 10−8 M to 10−4 M. The criteria of heterophyllous induction include the change in leaf morphology, the elongation of the petiole and the root, and the shortening of the internode. The measurements of the organs were taken after 3 weeks of treatment.
The responses are the opposite, i.e. the shortening of the petiole and the root, and the elongation of the internode.
The effects of these concentrations are not significantly different.
The nodes are clustered at the shoot apex, and the internode lengths were calculated by dividing the total length by the number of nodes.
Biphasic responses, with distinct optimal concentration in each phase.
Comparison of the effective concentrations of (+)-ABA and (−)-ABA on the induction of heterophyllous switch in M. quadrifolia
Organ | ABA isomer | Concentration (M) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Ineffective | Low dosage effecta | Effective range | Optimal | |||||||
| Petiole | (+) | 10−8 | 10−7 | 10−6∼10−4 | 10−5 | |||||
| (−) | – | – | 10−8∼10−4 | 10−7∼10−6b | ||||||
| Internode | (+) | 10−8 | 10−7 | 10−6∼10−4 | 10−5∼10−4b | |||||
| (−) | – | 10−8 | 10−7∼10−4 | ≥10−7c | ||||||
| Root | (+) | 10−8 | – | 10−7∼10−4 | Earlyd | Lated | ||||
| 10−5 | 10−6 | |||||||||
| (−) | – | – | 10−8∼10−4 | Earlyd | Lated | |||||
| 10−7∼10−6b | 10−5∼10−4b | |||||||||
Organ | ABA isomer | Concentration (M) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Ineffective | Low dosage effecta | Effective range | Optimal | |||||||
| Petiole | (+) | 10−8 | 10−7 | 10−6∼10−4 | 10−5 | |||||
| (−) | – | – | 10−8∼10−4 | 10−7∼10−6b | ||||||
| Internode | (+) | 10−8 | 10−7 | 10−6∼10−4 | 10−5∼10−4b | |||||
| (−) | – | 10−8 | 10−7∼10−4 | ≥10−7c | ||||||
| Root | (+) | 10−8 | – | 10−7∼10−4 | Earlyd | Lated | ||||
| 10−5 | 10−6 | |||||||||
| (−) | – | – | 10−8∼10−4 | Earlyd | Lated | |||||
| 10−7∼10−6b | 10−5∼10−4b | |||||||||
Plants were treated with the optically pure ABA enantiomer at various concentrations, ranging from 10−8 M to 10−4 M. The criteria of heterophyllous induction include the change in leaf morphology, the elongation of the petiole and the root, and the shortening of the internode. The measurements of the organs were taken after 3 weeks of treatment.
The responses are the opposite, i.e. the shortening of the petiole and the root, and the elongation of the internode.
The effects of these concentrations are not significantly different.
The nodes are clustered at the shoot apex, and the internode lengths were calculated by dividing the total length by the number of nodes.
Biphasic responses, with distinct optimal concentration in each phase.
Applied (−)-ABA accumulates in the heterophyllous responsive shoot apices
Feeding of labelled compounds and mass spectrometric studies were carried out to understand the correlation between growth response and ABA concentration in the heterophyllous responsive shoot apices. The scheme to synthesize optically pure (+)- and (−)- ABA analogues was developed previously, with deuterium atoms replacing the protons on the trans double bond of the side chain, positions which are not involved in known metabolism processes and are not exchangeable in the plant or the medium (Abrams et al., 2003). These compounds are useful for tracing the fate of exogenously applied ABA and also enable exogenous ABA to be distinguished from endogenous ABA. To correlate with the developmental studies, plants were supplied with 1 μM of either isomer of deuterated ABA, and analysed weekly for 3 weeks. Measurements of ABA and ABA metabolites are listed in Table 4. In addition, it was found that, after a week, the added ABA in the medium reached a relatively steady concentration, which persists during the 3-week period of the experiments: the contents of (+)-[4,5-d2]-ABA and (−)-[4,5-d2]-ABA in the medium were similar, 146±35 ng ml−1 (0.55 μM) and 138±16 ng ml−1 (0.52 μM), respectively. As expected, because the experiments were conducted in the light, the ABA in the medium equilibrated to a 1:1 mixture with 2-trans ABA.
Contents of ABA and ABA metabolites in heterophyllous responsive shoot apex tissues during a time-course of ABA treatmenta
ABA isomer | Concentration (nmol g−1 FW) | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Endogenous (d0) | Labelled (d2) | |||||||||||||
| ABA | PA | DPA | ABA | PA | DPA | 7′-OH ABA | ||||||||
| Week 1 | Control | 0.27±0.00 | n.d. | n.d. | ||||||||||
| (+) | 0.23±0.04 | n.d | 1.88±0.50 | 0.11±0.08 | n.d. | 14.54±5.04 | n.d. | |||||||
| (−) | 0.30±0.11 | 0.29±0.07 | 1.88±0.71 | 1.80±0.38 | n.d. | |||||||||
| Week 2 | Control | 0.23±0.04 | n.d. | n.d. | ||||||||||
| (+) | 0.15±0.00 | n.d. | 2.41±0.82 | 0.15±0.04 | 0.74±0.00 | 13.63±3.24 | n.d. | |||||||
| (−) | 0.19±0.08 | 0.04±0.00 | 4.01±0.85 | 2.74±0.94 | 0.82±0.53 | |||||||||
| Week 3 | Control | 0.11±0.00 | 0.18±0.11 | 0.78±0.32 | ||||||||||
| (+) | 0.23±0.08 | 0.11±0.04 | 2.87±0.50 | 1.20±0.19 | 0.04±0.04 | 12.39±1.58 | n.d. | |||||||
| (−) | 0.30±0.08 | 0.29±0.11 | 5.50±1.67 | 1.28±0.11 | 0.50±0.04 | |||||||||
ABA isomer | Concentration (nmol g−1 FW) | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Endogenous (d0) | Labelled (d2) | |||||||||||||
| ABA | PA | DPA | ABA | PA | DPA | 7′-OH ABA | ||||||||
| Week 1 | Control | 0.27±0.00 | n.d. | n.d. | ||||||||||
| (+) | 0.23±0.04 | n.d | 1.88±0.50 | 0.11±0.08 | n.d. | 14.54±5.04 | n.d. | |||||||
| (−) | 0.30±0.11 | 0.29±0.07 | 1.88±0.71 | 1.80±0.38 | n.d. | |||||||||
| Week 2 | Control | 0.23±0.04 | n.d. | n.d. | ||||||||||
| (+) | 0.15±0.00 | n.d. | 2.41±0.82 | 0.15±0.04 | 0.74±0.00 | 13.63±3.24 | n.d. | |||||||
| (−) | 0.19±0.08 | 0.04±0.00 | 4.01±0.85 | 2.74±0.94 | 0.82±0.53 | |||||||||
| Week 3 | Control | 0.11±0.00 | 0.18±0.11 | 0.78±0.32 | ||||||||||
| (+) | 0.23±0.08 | 0.11±0.04 | 2.87±0.50 | 1.20±0.19 | 0.04±0.04 | 12.39±1.58 | n.d. | |||||||
| (−) | 0.30±0.08 | 0.29±0.11 | 5.50±1.67 | 1.28±0.11 | 0.50±0.04 | |||||||||
Values are shown as mean ±SE of three replicates with eight shoot apices in each. n.d.: not detected.
Contents of ABA and ABA metabolites in heterophyllous responsive shoot apex tissues during a time-course of ABA treatmenta
ABA isomer | Concentration (nmol g−1 FW) | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Endogenous (d0) | Labelled (d2) | |||||||||||||
| ABA | PA | DPA | ABA | PA | DPA | 7′-OH ABA | ||||||||
| Week 1 | Control | 0.27±0.00 | n.d. | n.d. | ||||||||||
| (+) | 0.23±0.04 | n.d | 1.88±0.50 | 0.11±0.08 | n.d. | 14.54±5.04 | n.d. | |||||||
| (−) | 0.30±0.11 | 0.29±0.07 | 1.88±0.71 | 1.80±0.38 | n.d. | |||||||||
| Week 2 | Control | 0.23±0.04 | n.d. | n.d. | ||||||||||
| (+) | 0.15±0.00 | n.d. | 2.41±0.82 | 0.15±0.04 | 0.74±0.00 | 13.63±3.24 | n.d. | |||||||
| (−) | 0.19±0.08 | 0.04±0.00 | 4.01±0.85 | 2.74±0.94 | 0.82±0.53 | |||||||||
| Week 3 | Control | 0.11±0.00 | 0.18±0.11 | 0.78±0.32 | ||||||||||
| (+) | 0.23±0.08 | 0.11±0.04 | 2.87±0.50 | 1.20±0.19 | 0.04±0.04 | 12.39±1.58 | n.d. | |||||||
| (−) | 0.30±0.08 | 0.29±0.11 | 5.50±1.67 | 1.28±0.11 | 0.50±0.04 | |||||||||
ABA isomer | Concentration (nmol g−1 FW) | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Endogenous (d0) | Labelled (d2) | |||||||||||||
| ABA | PA | DPA | ABA | PA | DPA | 7′-OH ABA | ||||||||
| Week 1 | Control | 0.27±0.00 | n.d. | n.d. | ||||||||||
| (+) | 0.23±0.04 | n.d | 1.88±0.50 | 0.11±0.08 | n.d. | 14.54±5.04 | n.d. | |||||||
| (−) | 0.30±0.11 | 0.29±0.07 | 1.88±0.71 | 1.80±0.38 | n.d. | |||||||||
| Week 2 | Control | 0.23±0.04 | n.d. | n.d. | ||||||||||
| (+) | 0.15±0.00 | n.d. | 2.41±0.82 | 0.15±0.04 | 0.74±0.00 | 13.63±3.24 | n.d. | |||||||
| (−) | 0.19±0.08 | 0.04±0.00 | 4.01±0.85 | 2.74±0.94 | 0.82±0.53 | |||||||||
| Week 3 | Control | 0.11±0.00 | 0.18±0.11 | 0.78±0.32 | ||||||||||
| (+) | 0.23±0.08 | 0.11±0.04 | 2.87±0.50 | 1.20±0.19 | 0.04±0.04 | 12.39±1.58 | n.d. | |||||||
| (−) | 0.30±0.08 | 0.29±0.11 | 5.50±1.67 | 1.28±0.11 | 0.50±0.04 | |||||||||
Values are shown as mean ±SE of three replicates with eight shoot apices in each. n.d.: not detected.
In both treated and untreated plants, it was found that the concentration of endogenous ABA in the heterophyllous responsive shoot apex tissues was within a 2–3-fold difference, in the range of 0.1–0.3 nmol g−1 FW (Table 4). Paired t-tests comparing measured values of endogenous ABA between the experimental groups in each week and between the same groups in different weeks showed no significant differences (P <0.05, n=3) except in week 2 the value in (+)-ABA treated tissues was significantly lower than that of the control (mean difference 3.50), and in week 3 the values in both (+)- and (−)-ABA treated tissues were significantly higher than that of the control (mean difference 2.58 and 4.08, respectively). In the first two weeks, the concentration of applied (+)-[4,5-d2]-ABA in these tissues was in the concentration range of the endogenous ABA, and in the third week, it became 8–10-fold, reaching 1.2 nmol g−1 FW (mean difference 8.14, P <0.01, n=3). By contrast, starting in the first week, the concentrations of applied (−)-[4,5-d2]-ABA in the tissues were significantly higher than the concentrations of endogenous ABA (P <0.01, n=3), and the measured values ranged from 1.3–2.7 nmol g−1 FW during the 3-week period. This concentration is 4-fold to over 10-fold that of the endogenous ABA, and is more than 2–5-fold of the concentration of applied (−)-[4,5-d2]-ABA that remains in the medium. Thus the exogenously supplied (−)-ABA accumulates in the responsive shoot apex tissues.
It was also found that both treated and untreated tissues contain endogenous ABA metabolites, namely PA and DPA, while 7′-OH ABA and the ABA glucose ester were not detected. In the first and the second weeks, PA was detected only in (−)-ABA-treated shoot apices, whereas DPA was detectable in the tissues treated with either enantiomer but not in the control. In the third week, concentrations of endogenous ABA metabolites were significantly higher in (−)-ABA-treated tissues than in those treated with (+)-ABA (P <0.05, n=3). These results suggest that upon treatment, endogenous ABA was produced and further metabolized in the plant tissues. Moreover, for this stimulation of endogenous ABA synthesis, (−)-ABA has a stronger effect than (+)-ABA. In addition, in treated plants, derivatives of the applied ABA enantiomers were detected; (+)-[4,5-d2]-ABA was metabolized to PA and DPA, while (−)-[4,5-d2]-ABA was biotransformed to 7′-OH ABA (Table 4).
Discussion
Issues on ABA recognition
In M. quadrifolia the unnatural ABA enantiomer, (−)-ABA, not only induces a heterophyllous switch, but also produces stronger effects than the natural (+)-ABA. Like natural ABA, (−)-ABA causes growth responses that are progressive, reversible, dose-dependent, and organ-specific. Moreover, (−)-ABA is effective at lower concentrations than (+)-ABA, and the optimal concentration of (−)-ABA is also lower, by one to two orders of magnitude, depending on the organs. So far (−)-ABA has not been identified in plants; what has been seen in these experiments does not occur in nature. This raises the issue of ABA perception and how the action of (−)-ABA ties in with that of the natural (+)-ABA.
These data show that heterophyllous development has a distinct quantitative nature, and ABA affects individual morphogenic traits quantitatively. For each experiment reported here a set of plants was simultaneously treated with racemic ABA in order to effect a comparison. It was found that the extent of differential growth responses caused by racemic ABA, in every aspect, falls between those of the (+)-ABA and (−)-ABA (data not shown). Similar results have been reported in other systems (Smart et al., 1995). Dosage analyses further indicate that, for heterophyllous induction in M. quadrifolia, the optimal concentration and the effective range of (+)- and (−)-ABA differ with organs and developmental stages. This complexity associated with a developmental switch suggests a cell-autonomous nature of the ABA response.
The differential growth responses to ABA during heterophyllous induction include promoting effects, such as the elongation of roots and petioles, the formation of lateral roots, and the expansion of the leaf surface. Other aspects of the developmental switching include inhibitory effects, for example, shortening of internodes, senescence, and growth arrest. The effects of (−)-ABA are stronger than those of (+)-ABA in every parameter when individual organs are compared at specific developmental time points. However, if growth responses were measured as a sum of multiple tissues or over an accumulated period of time, different conclusions might result from comparing the effects of (+)- and (−)-ABA.
For example, regardless of the morphological change, when the growth response is compared using total leaf number, both (+)- and (−)-ABA would appear to be inhibitory at the end of the experimental period used here. Similarly, when the growth response is measured using total plant mass after 2 months of treatment, (−)-ABA would appear less effective than (+)-ABA in promoting growth or would have little effect compared with the untreated plants. These results further indicate that the heterophyllous switch consists of multiphasic developmental transition. Collectively, these observations provide useful reference for further investigations on the molecular mechanism of ABA recognition and signalling.
Endogenous versus exogenous ABA
The deuterated ABA analogues, used as standards for mass spectrometry and metabolism studies, allowed a distinction of the level and the fate of endogenous and exogenous ABA. Regardless of the treatment, endogenous ABA in heterophyllous-inducible shoot apices appeared to remain in a more or less constant range of 0.20±0.07 nmol g−1 FW, or approximately 0.2 μM. For switching the developmental path, (−)-ABA is effective at as low as 0.01 μM for both petioles and roots, and at 0.1 μM for the internodes, whereas (+)-ABA is effective when supplied at 1 μM for shoots and at 0.1 μM for roots. Following the induction via the treatment of 1 μM of either ABA enantiomer, the supplied ABA accumulates at 4–10-fold the concentration of endogenous ABA. These data indicate that both the exogenous ABA level and the internal tissue concentration of ABA associated with the developmental trigger are probably attainable by the tissues under normal growth conditions and, therefore, are within the physiological range.
The study of Smart et al. (1995) addressed similar questions on the role of (+)- and (−)-ABA on turion induction in the water plant Spirodela polyrrhiza L. These workers fed racemic, (+)- and (−)-ABA to the fronds and found that (−)-ABA had as strong or a stronger effect on growth inhibition and turion formation than the natural compound or the racemic mixture. This is comparable to what has been observed in M. quadrifolia. They measured natural ABA using an ELISA assay in fronds treated with (+)-ABA and were able to demonstrate that the concentrations of internal ABA required to induce turion formation was within a physiological range. The measurement of total ABA in fronds using high performance liquid chromatography/gas chromatography-electron capture detection showed that the concentrations of ABA [total of both (+)- and (−)-ABA] in the fronds treated with (−)-ABA were much higher than the ABA concentrations in the fronds treated with (+)-ABA. They concluded that the accumulation was due to the (−)-ABA enantiomer. A tight correlation has been consistently observed between the exogenous concentration of ABA and turion formation. However, with measurements in fronds, they could not find a simple internal concentration of ABA required for triggering the switch. This is possibly because the ABA-responsive tissue may represent only a fraction of the total plant material used for quantitative ABA analysis, as alluded to by these authors and shown to be the case for M. quadrifolia (Hsu et al., 2001).
It was found that supplied (−)-ABA accumulates in heterophyllous-responsive shoot apices at a higher concentration than the supplied (+)-ABA (Table 4). This may be because (−)-ABA is metabolized more slowly than (+)-ABA, as has been accounted for in cell suspension cultures of bromegrass and maize (Abrams et al., 1989; Balsevich et al., 1994a, b). Indeed, the concentration of the metabolite, 7′-OH ABA, represents only a fraction (approximately 30%) of the accumulated (−)-ABA. By contrast, metabolites of the supplied (+)-ABA exist in the tissues at 10-fold to over 100-fold the concentration of the hormone itself. Similarly, the metabolites of endogenous ABA are at high levels, approximately 10–20 times the concentration of endogenous ABA, which remains at a constant range (Table 4). However, the high levels of metabolites of (+)-ABA in tissues, both endogenous and exogenous, do not readily correlate with the physiological responses. Based on these observations, it is conceivable that either (−)-ABA itself or one of its metabolites, or both, can be recognized by the cellular component(s) of the signalling pathway and function similarly to (+)-ABA.
The role of ABA metabolism
In maize suspension cultures, (+)-ABA but not (−)-ABA has been shown to induce (+)-ABA hydroxylation (Cutler et al., 1997), and in potato and Arabidopsis both enantiomers induce ABA oxidation (Windsor and Zeevaart, 1997). The effect of inducing oxidation of ABA in M. quadrifolia would be to reduce internal, supplied and endogenous, (+)-ABA levels in (+)-ABA-treated tissues. If the M. quadrifolia ABA hydroxylation enzymes metabolize the natural (+)-ABA more rapidly than the (−)-ABA as occurs in maize and bromegrass (Abrams et al., 1989; Balsevich et al., 1994a, b), the (−)-ABA concentration would decline at a slower rate.
The authors have no data on the uptake and export of ABA enantiomers. However, it was found that applied ABA was metabolized in tissues of M. quadrifolia. On the other hand, the endogenous DPA concentration increases and accumulated in (−)-ABA-treated plants to higher levels than in those treated with (+)-ABA after 3 weeks. This indicates more endogenous ABA was synthesized in the (−)-ABA-treated tissues. Measurements of the metabolites and the hormone give a complete picture of the amount of ABA that had been synthesized and affected the system during the experiment. Thus the more persistent (−)-ABA causes increased biosynthesis of (+)-ABA that may contribute to triggering the switch at a lower concentration of (−)-ABA versus (+)-ABA.
In other biological responses elicited by (−)-ABA (Table 1) it is possible that the ABA-like effects observed are the results of increased ABA biosynthesis and catabolism. This may also be the case in the study on turion formation (Smart et al., 1995). In the effects on gene expression and growth inhibition in wheat embryos (Walker-Simmons et al., 1992), differential gene expression was observed while both enantiomers were equally effective germination inhibitors, indicating that (−)-ABA may have been acting by means other than by stimulating ABA biosynthesis. The effects of external stimuli such as supplied (+)- and (−)-ABA on endogenous hormone synthesis need to be considered for each individual system.
This work demonstrates that applied (−)-ABA stimulates ABA biosynthesis and suggests that it may contribute to at least part of the activity observed. Studies to block ABA biosynthesis using fluridone showed that either ABA enantiomer remained effective to induce the heterophyllous switch, despite the inhibitor's general toxicity to plant growth (H-J Wang, B-L Lin, unpublished results). Based on the findings in this work and the literature precedents, it cannot be ruled out that (−)-ABA itself is intrinsically active. It remains to be tested whether some of the metabolites also have a role for the induction. The work is now in progress.
Present address: National Health Research Institutes, 35 Keyan Road, Zhunan, Miaoli 350, Taiwan.
Abbreviations: ABA, abscisic acid; ABRH, ABA-responsive heterophylly; DPA, dihydrophaseic acid; LC-ES-MS-MS, liquid chromatography-electrospray ionization-tandem mass spectroscopy; PA, phaseic acid; 7′-OH ABA, 7′-hydroxy abscisic acid.
We are indebted to Su-Chuan Chiang for exceptional technical assistance. We thank Rung-Wu Chen and Hung-Chi Liu for help with the plant measurements, and Hung-Chi Liu also for help with preparing the figures and tables. We would also like to thank Dr Andrew Ross and Steve Ambrose for help with recording mass spectrometry data, as well as Ken Nelson for his technical assistance. This research was supported by the Cooperative Research Program of the National Science Council, Taiwan and the National Research Council, Canada, grant No. NSC 90-2311-B-169-002 and NSC 91-2311-B-169-002 to BLL. HJW is a recipient of a post-doctoral fellowship from National Science Council, Taiwan.
References
Abrams SR, Reaney MJT, Abrams GD, Mazurek T, Shaw AC, Gusta LV.
Abrams SR, Lamb N, Hill RD, Abrams GD, Yin XS, Fernets W, Laycock G, Gusta LV.
Abrams SR, Nelson K, Ambrose SJ.
Allsopp A.
Balsevich JJ, Abrams SR, Lamb N, König WA.
Balsevich JJ, Cutler AJ, Lamb N, Friesen LJ, Kurz EU, Perras MR, Abrams SR.
Bianco-Colomas J, Barthe P, Orlandini M, Le Page-Degivry MT.
Bommineni VR, Bethune TD, Hull KL, Dunstan DI.
Chandler JW, Abrams SR, Bartels D.
Chen J, Dellaporta S.
Cramer GR, Krishnan K, Abrams SR.
Cutler AJ, Squires TM, Loewen MK, Balsevich JJ.
Cummins WR, Sondheimer E.
Cummins WR, Kende H, Raschke K.
Deschamp PA, Cooke TJ.
Deschamp PA, Cooke TJ.
Dong J-Z, Dunstan DI.
Dong J-Z, Dunstan DI.
Dong J-Z, Pilate G, Abrams SR, Dunstan DI.
Dunstan DI, Bock CA, Abrams GD, Abrams SR.
Feurtado JA, Ambrose SJ, Cutler AJ, Ross ARS, Abrams SR, Kermode AR.
Gaudet JJ.
Gaudet JJ.
Gifford EM, Foster AS.
Goliber TE.
Goliber TE, Feldman LJ.
Gusta LV, Ewan B, Reaney MJT, Abrams SR.
Hampson CR, Reaney MJT, Abrams GD, Abrams SR, Gusta LV.
Hays DB, Rose P, Abrams SR, Moloney MM.
Hill RD, Liu JH, Durnin D, Lamb N, Shaw A, Abrams SR.
Hsu TC, Liu HC, Wang JS, Chen RW, Wang YC, Lin BL.
Jeannette E, Rona JP, Bardat F, Cornel D, Sotta B, Miginiac E.
Jepson I, Bray J, Jenkins G, Schuch W, Edwards K.
Jiang L, Abrams SR, Kermode AR.
Laetsch WM, Briggs WR.
Lin BL, Yang WJ.
Liu BLL.
Milborrow BV.
Nakano S, Todoroki Y, Hirai N, Ohigashi H.
Nambara E, Suzuki M, Abrams SR, McCarty DR, Kamiya Y, McCourt P.
Perras MR, Abrams SR, Balsevich JJ.
Robertson AJ, Reaney MJT, Wilen RW, Lamb N, Abrams SR, Gusta LV.
Rose PA, Lei B, Shaw AC, Barton DL, Walker-Simmons MK, Abrams SR.
Ross ARS, Ambrose SJ, Cutler AJ, Feurtado JA, Kermode AR, Nelson K, Zhou R, Abrams, SR.
Sambrook J, Fritsch EF, Maniatis T.
Sauter A, Abrams SR, Hartung W.
Schmitz N, Abrams SR, Kermode AR.
Shen J, Niwa T, Marumo S.
Smart CC, Fleming AJ, Chaloupkova K, Hanke DE.
Sondheimer E, Galson EC, Chang YP, Walton DC.
Todoroki Y, Hirai N, Koshimizu K.
Todoroki Y, Hirai N, Koshimizu K.
Todoroki Y, Hirai N, Koshimizu K.
Toorop PE, Bewley JD, Abrams SR, Hilhorst HWM.
Trewavas AJ, Jones HG.
Walker-Simmons MK, Anderberg RJ, Rose PA, Abrams SR.
Wilen RW, Fu P, Robertson AJ, Abrams SR, Low NH, Gusta LV.
Wilen RW, Hays DB, Mandel RM, Abrams SR, Moloney MM.
Wilmer JA, Abrams SR, Helsper JPFG, van der Plas LHW.
Windsor ML, Zeevaart JAD.
Windsor ML, Milborrow BV, Abrams SR.
Zaharia LI, Galka MM, Ambrose SJ, Abram SR.
Zeevaart JAD, Creelman RA.
Zhou R, Cutler AJ, Ambrose SJ, et al.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Comments