Rootstock-Mediated Effects on Cabernet Sauvignon Performance: Vine Growth, Berry Ripening, Flavonoids, and Aromatic Profiles

 ,
, 
Abstract
:1. Introduction
2. Results and Discussion
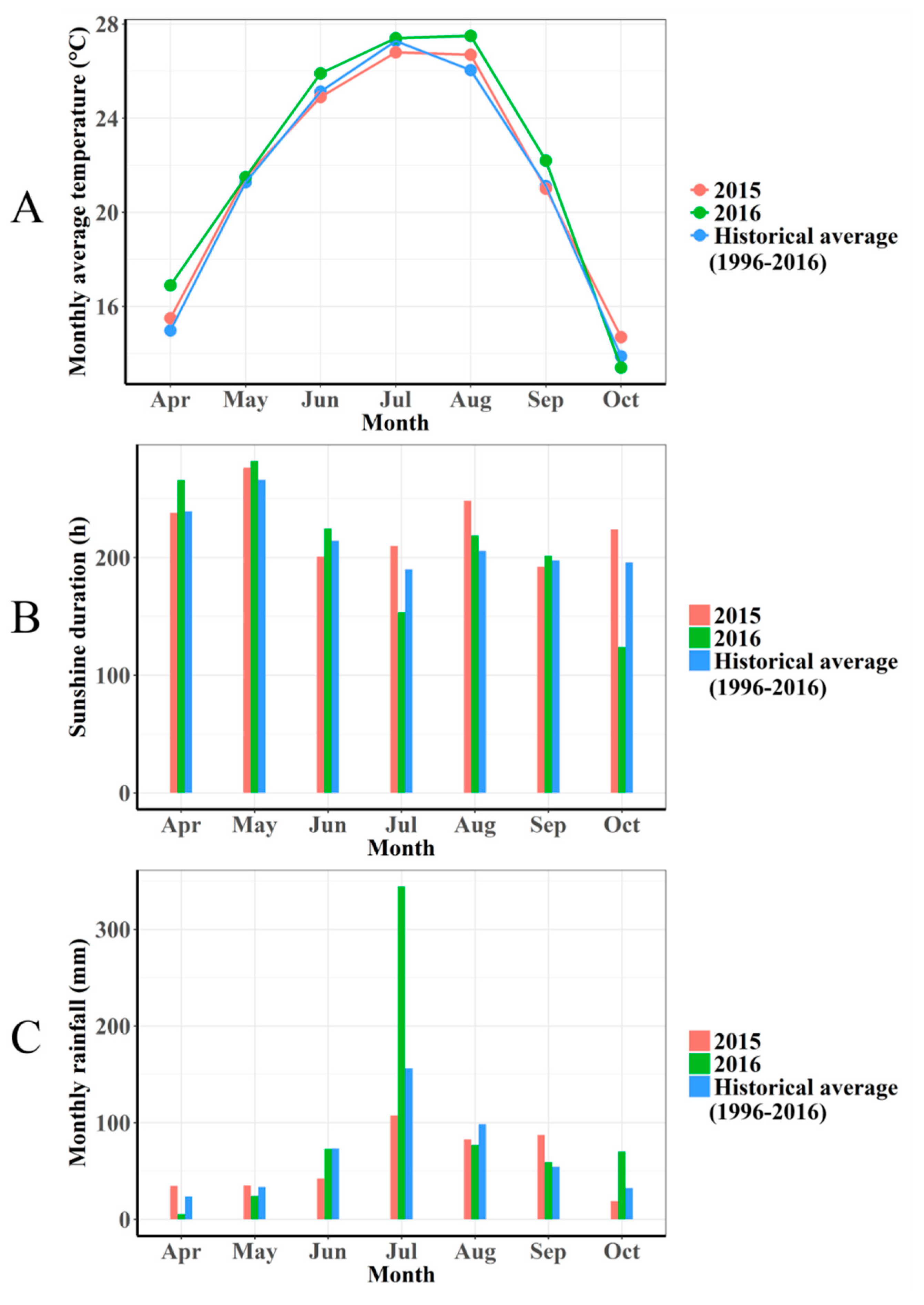
2.1. Meteorological Data
2.2. Effects of Rootstocks on Vine Growth
2.3. Effects of Rootstocks on Berry Physiochemical Parameters
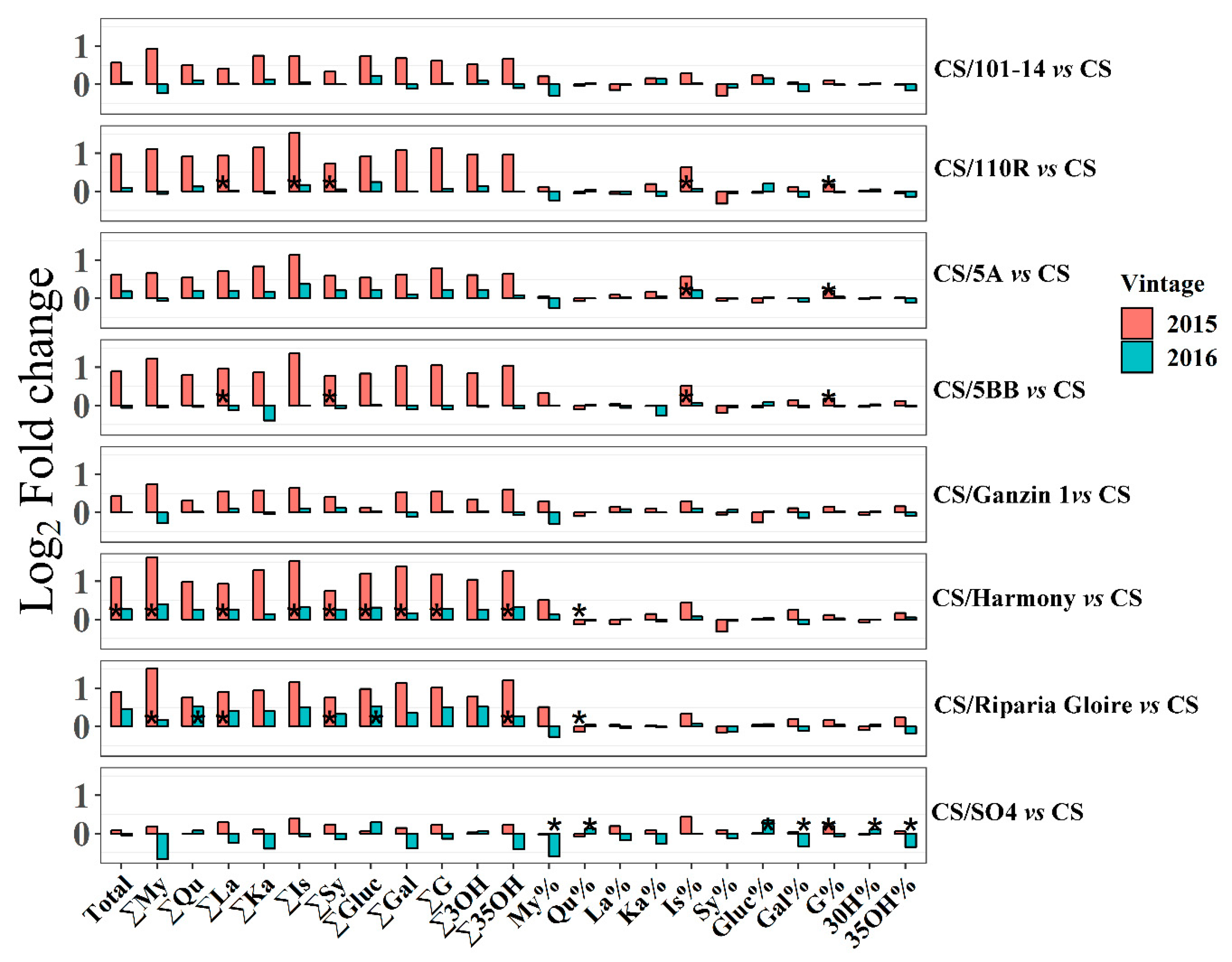
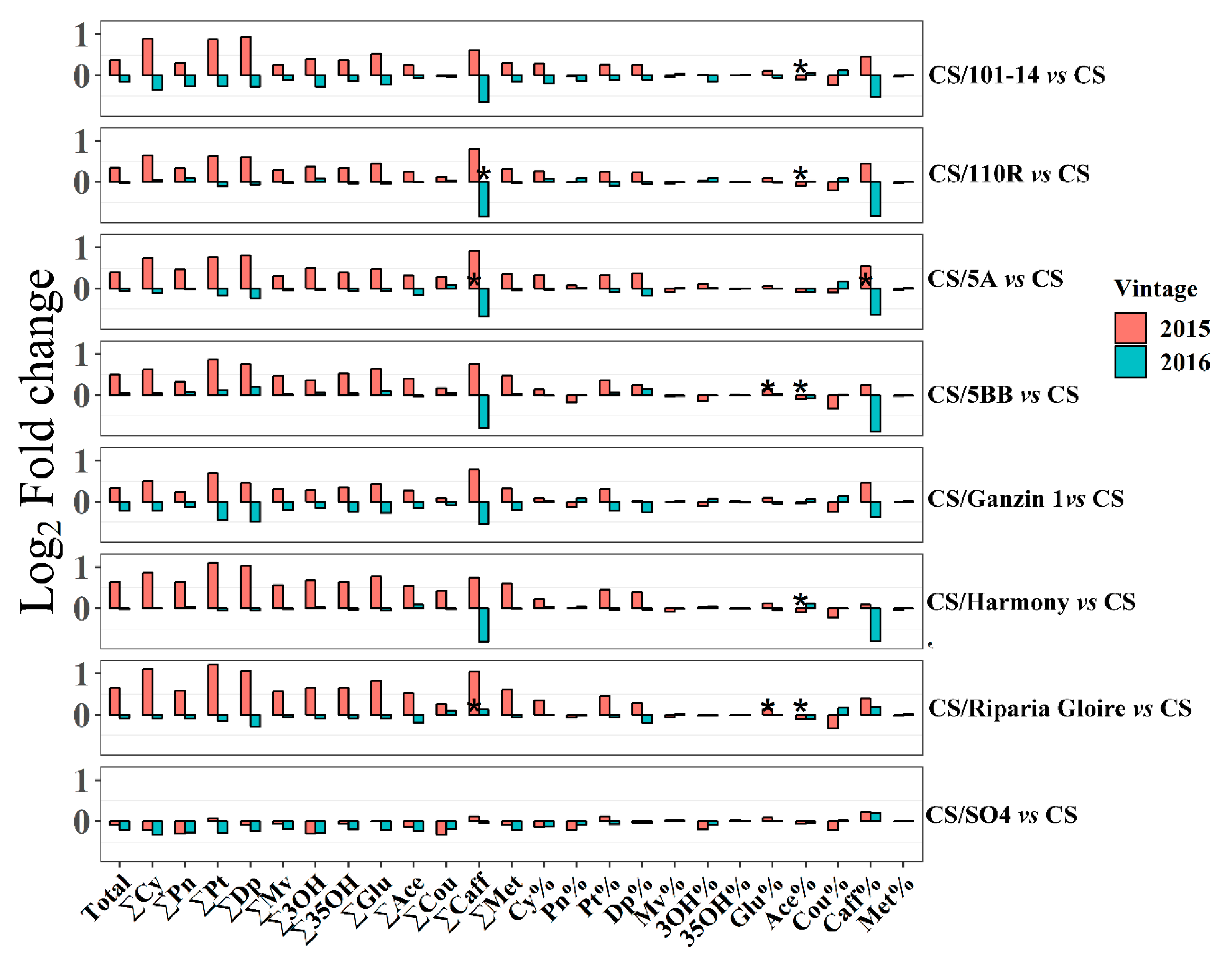
2.4. Effects of Rootstocks on Berry Flavonoids
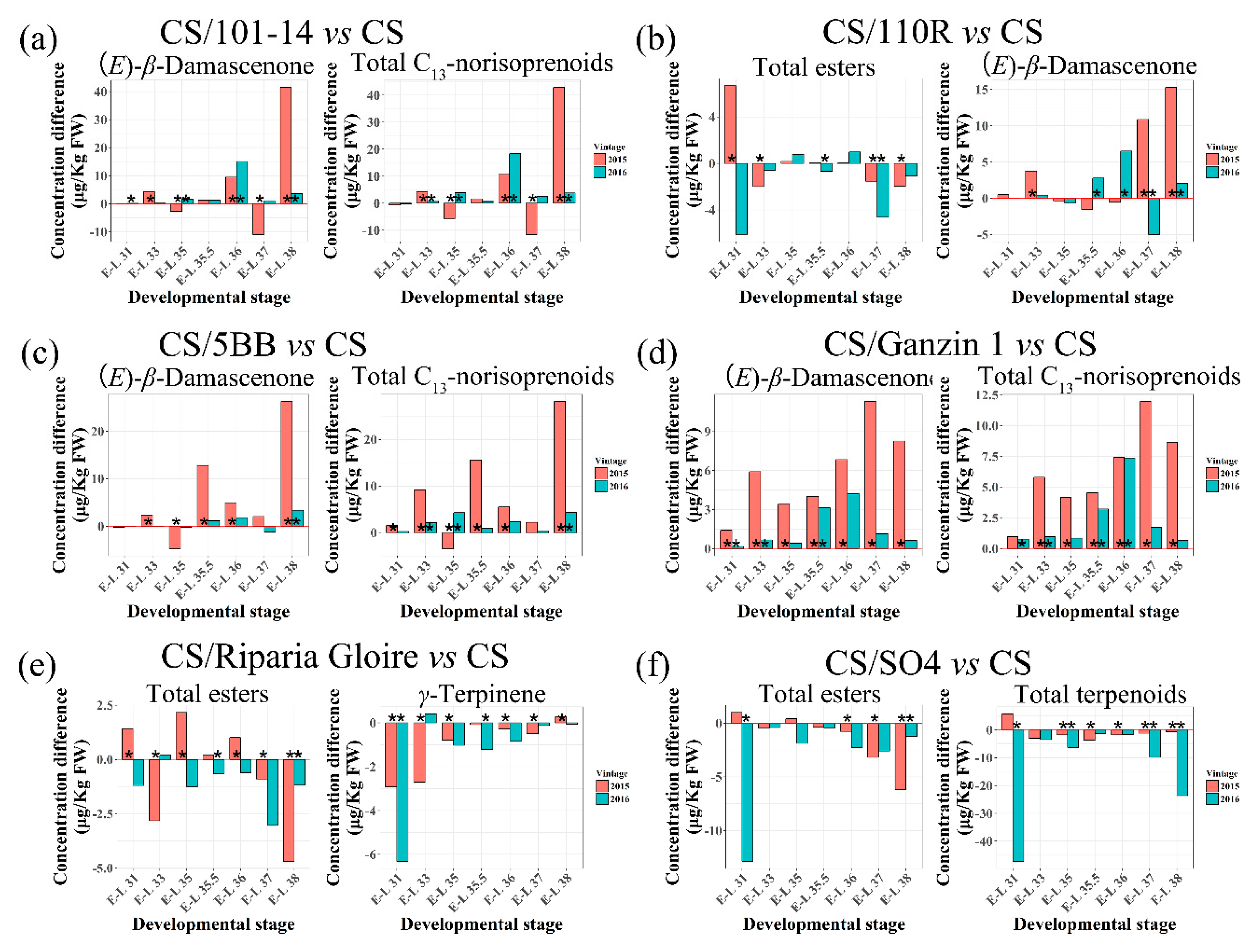
2.5. Effects of Rootstocks on Berry Volatile Compounds
3. Materials and Methods
3.1. Vineyard, Experiment Design, and Vine Management
3.2. Experiment Design and Sample Collection
3.3. Measurement of Vine Growth Parameters
3.4. Determinations of Berry Physiochemical Parameter
3.5. The Extraction of Flavonoids
3.6. High Performance Liquid Chromatography-Mass Spectrometry (HPLC-MS) Analysis of Flavonoids in Berry Skins
3.7. Extraction of Volatile Compounds
3.8. Gas Chromatography-Mass Spectrometry (GC-MS) Analysis of Volatile Compounds
3.9. Chemicals
3.10. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| O2PLS-DA | Bidirectional orthogonal partial least squares-discriminant analysis |
| VIP | Variable importance in projection |
| CS | Cabernet Sauvignon |
| M-VSP | Modified vertical shoot positioning |
| TSS | Total soluble solids |
| HPLC-MS | High performance liquid chromatography-mass spectrometry |
| C | (+)-Catechin |
| EC | (−)-Epicatechin |
| EGC | (−)-Epigallocatechin |
| ECG | (−)-Epicatechin-3-O-galate |
| HS-SPME | Headspace solid phase microextraction |
| GC-MS | Gas chromatography-mass spectrometry |
| RI | Retention indices |
| AMDIS | Automated MassSpectral Deconvolution and Identification System |
References
- Ezzahouani, A.; Williams, L.E. The influence of rootstock on leaf water potential, yield, and berry composition of Ruby Seedless grapevines. Am. J. Enol. Vitic. 1995, 46, 559–563. [Google Scholar]
- Avenant, E.; Avenant, J.H.; Barnard, R.O. The effect of three rootstock cultivars, potassium soil applications and foliar sprays on yield and quality of Vitis vinifera L. cv. Ronelle in South Africa. S. Afr. J. Enol. Vitic. 1997, 18, 31–38. [Google Scholar] [CrossRef]
- Stevens, R.M.; Pech, J.M.; Gibberd, M.R.; Walker, R.R.; Jones, J.A.; Taylor, J.; Nicholas, P.R. Effect of reduced irrigation on growth, yield, ripening rates and water relations of Chardonnay vines grafted to five rootstocks. Aust. J. Grape Wine Res. 2008, 14, 177–190. [Google Scholar] [CrossRef]
- Koundouras, S.; Hatzidimitriou, E.; Karamolegkou, M.; Dimopoulou, E.; Kallithraka, S.; Tsialtas, J.T.; Zioziou, E.; Nikolaou, N.; Kotseridis, Y. Irrigation and rootstock effects on the phenolic concentration and aroma potential of Vitis vinifera L. cv. Cabernet Sauvignon grapes. J. Agric. Food Chem. 2009, 57, 7805–7813. [Google Scholar] [CrossRef] [PubMed]
- Wooldridge, J.; Louw, P.J.E.; Conradie, W.J. Effects of rootstock on grapevine performance, petiole and must composition, and overall wine score of Vitis vinifera cv. Chardonnay and Pinot noir. S. Afr. J. Enol. Vitic. 2010, 31, 45–48. [Google Scholar] [CrossRef]
- Chitarra, W.; Perrone, I.; Avanzato, C.G.; Minio, A.; Boccacci, P.; Santini, D.; Gilardi, G.; Siciliano, I.; Gullino, M.L.; Delledonne, M.; et al. Grapevine grafting: Scion transcript profiling and defense-related metabolites induced by rootstocks. Front. Plant Sci. 2017, 8, 654. [Google Scholar] [CrossRef]
- Jogaiah, S.; Oulkar, D.P.; Banerjee, K.; Sharma, J.; Patil, A.G.; Maske, S.R.; Somkuwar, R.G. Biochemically induced variations during some phenological stages in Thompson Seedless grapevines grafted on different rootstocks. S. Afr. J. Enol. Vitic. 2013, 34, 36–45. [Google Scholar] [CrossRef]
- Cookson, S.J.; Ollat, N. Grafting with rootstocks induces extensive transcriptional re-programming in the shoot apical meristem of grapevine. BMC Plant Biol. 2013, 13, 147. [Google Scholar] [CrossRef]
- Berdeja, M.; Hilbert, G.; Dai, Z.; Lafontaine, M.; Stoll, M.; Schultz, H.; Delrot, S. Effect of water stress and rootstock genotype on Pinot Noir berry composition. Aust. J. Grape Wine Res. 2014, 20, 409–421. [Google Scholar] [CrossRef]
- Nelson, C.C.; Kennedy, J.A.; Zhang, Y.; Kurtural, S.K. Applied water and rootstocks affect productivity and anthocyanin composition of Zinfandel in central California. Am. J. Enol. Vitic. 2016, 67, 18–28. [Google Scholar] [CrossRef]
- Keller, M.; Mills, L.J.; Harbertson, J.F. Rootstock effects on deficit-irrigated winegrapes in a dry climate: Vigor, yield formation, and fruit ripening. Am. J. Enol. Vitic. 2012, 63, 29–39. [Google Scholar] [CrossRef]
- Conradie, W.J. Liming and choice of rootstocks as cultural techniques for vines in acid soils. S. Afr. J. Enol. Vitic. 1983, 4, 39–44. [Google Scholar] [CrossRef]
- Wooldridge, J.; Olivier, M.P. Effects of weathered soil parent materials on Merlot grapevines grafted onto 110 Richter and 101-14Mgt rootstocks. S. Afr. J. Enol. Vitic. 2014, 35, 59–67. [Google Scholar] [CrossRef]
- Sommer, K.; Clingeleffer, P.; Ollat, N. Effects of minimal pruning on grapevine canopy development, physiology and cropping level in both cool and warm climates. Wein-Wissenschaft 1993, 48, 135–139. [Google Scholar]
- Benz, M.J.; Anderson, M.M.; Williams, M.A.; Wolpert, J.A. Viticultural performance of three Malbec clones on two rootstocks in Oakville, Napa Valley, California. Am. J. Enol. Vitic. 2007, 58, 262–267. [Google Scholar]
- Pulko, B.; Vršič, S.; Valdhuber, J. Influence of various rootstocks on the yield and grape composition of Sauvignon blanc. Czech J. Food Sci. 2012, 30, 467–473. [Google Scholar] [CrossRef]
- Loureiro, M.D.; Moreno-Sanz, P.; García, A.; Fernández, O.; Fernández, N.; Suárez, B. Influence of rootstock on the performance of the Albarin Negro minority grapevine cultivar. Sci. Hortic. 2016, 201, 145–152. [Google Scholar] [CrossRef]
- Corso, M.; Vannozzi, A.; Ziliotto, F.; Zouine, M.; Maza, E.; Nicolato, T.; Vitulo, N.; Meggio, F.; Valle, G.; Bouzayen, M.; et al. Grapevine rootstocks differentially affect the rate of ripening and modulate auxin-related genes in Cabernet Sauvignon berries. Front. Plant Sci. 2016, 7, 14. [Google Scholar] [CrossRef]
- Jin, Z.-X.; Sun, T.-Y.; Sun, H.; Yue, Q.-Y.; Yao, Y.-X. Modifications of ‘Summer Black’grape berry quality as affected by the different rootstocks. Sci. Hortic. 2016, 210, 130–137. [Google Scholar] [CrossRef]
- Jin, Z.X.; Sun, H.; Sun, T.Y.; Wang, Q.J.; Yao, Y.X. Modifications of ‘Gold Finger’ grape berry quality as affected by the different rootstocks. J. Agric. Food Chem. 2016, 64, 4189–4197. [Google Scholar] [CrossRef]
- Warschefsky, E.J.; Klein, L.L.; Frank, M.H.; Chitwood, D.H.; Londo, J.P.; von Wettberg, E.J.B.; Miller, A.J. Rootstocks: Diversity, domestication, and impacts on shoot phenotypes. Trends Plant Sci. 2016, 21, 418–437. [Google Scholar] [CrossRef]
- Habran, A.; Commisso, M.; Helwi, P.; Hilbert, G.; Negri, S.; Ollat, N.; Gomès, E.; van Leeuwen, C.; Guzzo, F.; Delrot, S. Roostocks/scion/nitrogen interactions affect secondary metabolism in the grape berry. Front. Plant Sci. 2016, 7, 1134. [Google Scholar] [CrossRef] [PubMed]
- Nuzzo, V.; Matthews, M.A. Response of fruit growth and ripening to crop level in dry-farmed Cabernet Sauvignon on four rootstocks. Am. J. Enol. Vitic. 2006, 57, 314–324. [Google Scholar]
- Wolpert, J.A.; Smart, D.R.; Anderson, M. Lower petiole potassium concentration at bloom in rootstocks with Vitis berlandieri genetic backgrounds. Am. J. Enol. Vitic. 2005, 56, 163–169. [Google Scholar]
- Koblet, W.; Candolfivasconcelos, M.C.; Zweifel, W.; Howell, G.S. Influence of leaf removal, rootstock and trainning system on yield and fruit composition of Pinot Noir grapevines. Am. J. Enol. Vitic. 1994, 45, 181–187. [Google Scholar]
- Kodur, S.; Tisdall, J.M.; Clingeleffer, P.R.; Walker, R.R. Regulation of berry quality parameters in ‘Shiraz’ grapevines through rootstocks (Vitis). Vitis 2013, 52, 125–128. [Google Scholar]
- Suriano, S.; Alba, V.; Di Gennaro, D.; Suriano, M.S.; Savino, M.; Tarricone, L. Genotype/rootstocks effect on the expression of anthocyanins and flavans in grapes and wines of Greco Nero n. (Vitis vinifera L.). Sci. Hortic. 2016, 209, 309–315. [Google Scholar] [CrossRef]
- Garcia, M.; Gallego, P.; Daverède, C.; Ibrahim, H. Effect of three rootstocks on grapevine (Vitis vinifera L.) cv. Negrette, grown hydroponically. I. Potassium, calcium and magnesium nutrition. S. Afr. J. Enol. Vitic. 2001, 22, 101–103. [Google Scholar] [CrossRef]
- Brancadoro, L.; Valenti, L.; Reina, A.; Scienza, A. Potassium content of grapevine during the vegetative period-the role of the rootstock. J. Plant Nutr. 1994, 17, 2165–2175. [Google Scholar] [CrossRef]
- Harbertson, J.F.; Keller, M. Rootstock effects on deficit-irrigated winegrapes in a dry climate: Grape and wine composition. Am. J. Enol. Vitic. 2012, 63, 40–48. [Google Scholar] [CrossRef]
- Gastón, G.; Encarna, G.; Bautista-Ortín, A.B.; Garde-Cerdán, T.; Moreno-Simunovic, Y.; Martínez-Gil, A.M. Rootstock effects on grape anthocyanins, skin and seed proanthocyanidins and wine color and phenolic compounds from Vitis vinifera L. Merlot grapevines. J. Sci. Food Agric. 2019. [CrossRef]
- Olarte Mantilla, S.M.; Collins, C.; Iland, P.G.; Kidman, C.M.; Ristic, R.; Boss, P.K.; Jordans, C.; Bastian, S.E.P. Shiraz (Vitis vinifera L.) berry and wine sensory profiles and composition are modulated by rootstocks. Am. J. Enol. Vitic. 2018, 69, 32–44. [Google Scholar] [CrossRef]
- Gollop, R.; Farhi, S.; Perl, A. Regulation of the leucoanthocyanidin dioxygenase gene expression in Vitis vinifera. Plant Sci. 2001, 161, 579–588. [Google Scholar] [CrossRef]
- Wang, Y.; He, Y.N.; Chen, W.K.; He, F.; Chen, W.; Cai, X.D.; Duan, C.Q.; Wang, J. Effects of cluster thinning on vine photosynthesis, berry ripeness and flavonoid composition of Cabernet Sauvignon. Food Chem. 2018, 248, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Coombe, B. Growth stages of the grapevine: Adoption of a system for identifying grapevine growth stages. Aust. J. Grape Wine Res. 1995, 1, 104–110. [Google Scholar] [CrossRef]
- Liang, N.-N.; He, F.; Pan, Q.-H.; Wang, J.; Reeves, M.J.; Duan, C.-Q. Optimization of sample preparation and phloroglucinol analysis of Marselan grape skin proanthocyanidins using HPLC-DAD-ESI-MS/MS. S. Afr. J. Enol. Vitic. 2012, 33, 122–131. [Google Scholar]
- Downey, M.O.; Mazza, M.; Krstic, M.P. Development of a stable extract for anthocyanins and flavonols from grape skin. Am. J. Enol. Vitic. 2007, 58, 358–364. [Google Scholar]
- Li, S.-Y.; He, F.; Zhu, B.-Q.; Wang, J.; Duan, C.-Q. Comparison of phenolic and chromatic characteristics of dry red wines made from native Chinese grape species and Vitis Vinifera. Int. J. Food Prop. 2016, 20, 2134–2146. [Google Scholar] [CrossRef]
- Sun, R.-Z.; Cheng, G.; Li, Q.; He, Y.-N.; Wang, Y.; Lan, Y.-B.; Li, S.-Y.; Zhu, Y.-R.; Song, W.-F.; Zhang, X.; et al. Light-induced variation in phenolic compounds in Cabernet Sauvignon Grapes (Vitis vinifera L.) involves extensive transcriptome reprogramming of biosynthetic enzymes, transcription factors, and phytohormonal regulators. Front. Plant Sci. 2017, 8, 547. [Google Scholar] [CrossRef] [PubMed]
- He, J.-J.; Liu, Y.-X.; Pan, Q.-H.; Cui, X.-Y.; Duan, C.-Q. Different anthocyanin profiles of the skin and the pulp of Yan73 (Muscat Hamburg × Alicante Bouschet) grape berries. Molecules 2010, 15, 1141–1153. [Google Scholar] [CrossRef]
- Lan, Y.-B.; Qian, X.; Yang, Z.-J.; Xiang, X.-F.; Yang, W.-X.; Liu, T.; Zhu, B.-Q.; Pan, Q.-H.; Duan, C.-Q. Striking changes in volatile profiles at sub-zero temperatures during over-ripening of ‘Beibinghong’grapes in Northeastern China. Food Chem. 2016, 212, 172–182. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Variation | Stock/Scion Diameter | Estimated Yield (kg/Vine) | Pruning Weight (kg/Vine) | Crop Load (Yield/Pruning Weight) |
|---|---|---|---|---|
| Rootstock (R) | ||||
| CS | 1.00 a a | 4.27 | 1.26 | 4.26 |
| CS/101-14 | 0.78 bc | 3.38 | 1.41 | 3.02 |
| CS/110R | 0.75 bcd | 3.52 | 1.38 | 3.08 |
| CS/5A | 0.77 bcd | 4.10 | 1.40 | 4.15 |
| CS/5BB | 0.75 bcd | 3.28 | 1.36 | 2.79 |
| CS/Ganzin 1 | 0.69 cd | 3.47 | 1.40 | 3.11 |
| CS/Harmony | 0.83 b | 3.08 | 1.29 | 3.30 |
| CS/Riparia Gloire | 0.67 d | 3.79 | 1.29 | 3.91 |
| CS/SO4 | 0.71 cd | 3.92 | 1.48 | 3.21 |
| Vintage (V) | ||||
| 2015 | 0.77 | 2.26 b | 1.79 b | 1.30 b |
| 2016 | 0.77 | 5.03 a | 0.93 a | 5.55 a |
| Significanceb | ||||
| R × S | ns c | ns | ns | ns |
| Source of Variation | Berry Weight (g/100 Berries) | Total Soluble Solids (oBrix) | Titratable Acidity (g/L) | pH |
|---|---|---|---|---|
| Rootstock (R) | ||||
| CS | 133.82 a | 20.10 ab | 5.55 | 3.50 |
| CS/101-14 | 133.64 | 19.73 ab | 6.44 | 3.38 |
| CS/110R | 127.52 | 20.05 ab | 5.48 | 3.39 |
| CS/5A | 144.87 | 19.35 ab | 5.12 | 3.35 |
| CS/5BB | 137.27 | 19.65 ab | 6.01 | 3.35 |
| CS/Ganzin 1 | 153.33 | 20.1 ab | 5.39 | 3.56 |
| CS/Harmony | 134.17 | 20.55 a | 5.18 | 3.46 |
| CS/Riparia Gloire | 142.64 | 19.6 ab | 4.68 | 3.42 |
| CS/SO4 | 130.00 | 18.28 b | 6.11 | 3.42 |
| Vintage (V) | ||||
| 2015 | 123.74 b | 20.24 a | 5.97 | 3.46 |
| 2016 | 147.62 a | 19.08 b | 5.39 | 3.38 |
| Significanceb | ||||
| R × S | ns c | ns | ns | ns |
| O2PLS-DA Models | Components a | R2X (cum) | R2Y (cum) | Q2 (cum) | pCV-ANOVA | 500 Permutation Tests | |
|---|---|---|---|---|---|---|---|
| R2Y Intercept | Q2Y Intercept | ||||||
| CS/101-14 vs. CS | 1 + 6 | 0.94 | 0.78 | 0.61 | 4.31 × 10−5 | 0.39 | −0.67 |
| CS/110R vs. CS | 1 + 5 | 0.91 | 0.86 | 0.72 | 5.07 × 10−10 | 0.36 | −0.63 |
| CS/5A vs. CS | 1 + 2 | 0.92 | 0.52 | 0.38 | 0.09 | 0.20 | −0.29 |
| CS/5BB vs. CS | 1 + 7 | 0.92 | 0.90 | 0.81 | 3.35 × 10−13 | 0.43 | −0.83 |
| CS/Ganzin 1 vs. CS | 1 + 6 | 0.93 | 0.84 | 0.74 | 1.00 × 10−9 | 0.40 | −0.71 |
| CS/Harmony vs. CS | 1 + 2 | 0.93 | 0.51 | 0.32 | 0.28 | 0.21 | −0.30 |
| CS/Riparia Gloire vs. CS | 1 + 8 | 0.93 | 0.91 | 0.68 | 4.03 × 10−7 | 0.48 | −0.99 |
| CS/SO4 vs. CS | 1 + 6 | 0.94 | 0.94 | 0.87 | 1.46 × 10−14 | 0.41 | −0.81 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Chen, W.-K.; Gao, X.-T.; He, L.; Yang, X.-H.; He, F.; Duan, C.-Q.; Wang, J. Rootstock-Mediated Effects on Cabernet Sauvignon Performance: Vine Growth, Berry Ripening, Flavonoids, and Aromatic Profiles. Int. J. Mol. Sci. 2019, 20, 401. https://doi.org/10.3390/ijms20020401
Wang Y, Chen W-K, Gao X-T, He L, Yang X-H, He F, Duan C-Q, Wang J. Rootstock-Mediated Effects on Cabernet Sauvignon Performance: Vine Growth, Berry Ripening, Flavonoids, and Aromatic Profiles. International Journal of Molecular Sciences. 2019; 20(2):401. https://doi.org/10.3390/ijms20020401
Chicago/Turabian StyleWang, Yu, Wei-Kai Chen, Xiao-Tong Gao, Lei He, Xiao-Hui Yang, Fei He, Chang-Qing Duan, and Jun Wang. 2019. "Rootstock-Mediated Effects on Cabernet Sauvignon Performance: Vine Growth, Berry Ripening, Flavonoids, and Aromatic Profiles" International Journal of Molecular Sciences 20, no. 2: 401. https://doi.org/10.3390/ijms20020401