


Abstract
Chardonnay Musqué vines were subjected to six crop levels (nonthinned control and cluster thinned at late bloom/early set, early stage I of berry growth, late stage I, lag phase, veraison) over four years (1999 to 2002). Wines were made from these treatments using yeast strain VL1 (Saccharomyces cerevisiae). Additional fruit from nonthinned vines was subjected to several enological treatments: EC1118 strain (S. bayanus) ± pectinase preparation Cinn-Free and VL3 strain (1999) and EC1118 ± Cinn-Free and AR2000 enzymes, and CY3079, D47, and VL3 strains (2000 and 2001). Enological treatments were compared chemically and sensorially and to the viticultural treatments. Thinning decreased yields and clusters per vine regardless of timing. Both berry and cluster weights decreased as time of thinning was delayed, suggesting yield compensation. Thinning at/after early stage I led to higher Brix, pH, and potentially volatile terpenes (PVT) relative to the control; titratable acidity (TA) decreased with later thinning; and free volatile terpenes (FVT) increased in some thinning treatments. There were substantial differences in wine pH, TA, and PVT among the different yeast strains and enzyme treatments. Use of prepress Cinn-Free led to increased TA and FVT, resulting from extraction of organic acids and terpenes from berry skins. VL1 increased wine TA and pH. VL1 and VL3 reduced wine PVT, perhaps because of enhanced β-glucosidase activity and/or biosynthesis/biotransformation of terpenes. Despite chemical composition differences among viticultural treatments, wines differed little sensorially. However, sensory differences did occur among yeast treatments, with VL1 wines being distinctive. Several sensory attributes that distinguished between yeast treatment wines were correlated with PVT.
Cluster thinning is commonly used as a corrective viticultural measure to improve fruit composition. Previous work focused on its role in altering grape and/or must composition, particularly common maturity indices such as Brix, pH, and titratable acidity (Petrie et al. 2000, Reynolds 1989, Reynolds et al. 1994a, Vasconcelos and Castagnoli 2000).
Monoterpenes are a chemical class of compounds present in many highly aromatic Vitis vinifera cultivars and can impart distinctive aromas and flavors to those cultivars belonging to the muscat family. Terpenes have been observed to be responsive to modifications to crop level (McCarthy 1986, Reynolds and Wardle 1989a, Reynolds et al. 1994a). Existing in the berry in free and glycosidically bound forms, the free monoterpenes contribute directly to wine aroma and flavor while the bound forms are rendered odorless. Upon chemical or enzymatic hydrolysis, glycosidically bound terpenes can be released from their sugar moiety, representing a pool of potential aroma and flavor. In most situations, changes in terpene concentration of berries and musts through cluster thinning have been associated with favorable modifications in flavor and/or aroma attributes (Reynolds and Wardle 1997, Reynolds et al. 1994b).
A specific group of Chardonnay clones, called Chardonnay Musqué (clones 77 and 809) (MAPA 1994), is popular in the Niagara region and is known for floral muscat-like aromas that distinguish the group from other more neutral clones. The accumulation of monoterpenes during fruit maturation is likely crucial to the identity and varietal character of Chardonnay Musqué. Viticultural and enological treatments that might increase monoterpene concentration of Chardonnay Musqué berries, musts, and wines are therefore of critical interest to winemakers who want to maximize expression of varietal character.
Yeast strain and enzyme manufacturers often claim that varietal typicity of certain wine cultivars may be improved through specific yeast strains and/or enzymes. Mention is made of β-glucosidase activity that allows a specific yeast strain or enzyme product to hydrolyze glycosidically bound terpenes to their free forms. The contribution of different yeast strains toward individual properties of wine has been examined (Antonelli et al. 1999, Dumont and Dulau 1996, Reynolds et al. 2001), and some commercial enzyme preparations have been marketed as having an ability to increase volatile characteristics of wine through β-glycosidase side activity (Canal-Llaubères 1994, Francis et al. 1996, Grossmann and Rapp 1988, Hesford et al. 1996).
Cluster thinning is potentially an expensive process in terms of labor and lost yield. If wine varietal typicity could be increased by a greater magnitude than cluster thinning in the winery through less expensive practices such as judicious yeast strain selection and/or enzyme use, then there would be potential significant cost saving. Our trial sought to examine the effect of cluster thinning applied to Chardonnay Musqué at five different phenological stages in terms of its impact on yield, vine performance, and berry, must, and wine composition. A parallel trial tested three yeast strains and enzymes that could be compared against each other and against viticultural treatments (Reynolds et al. 2007). The overall objective of this study was to determine the relative magnitude of effects of viticultural and enological treatments upon wine composition and sensory attributes of Chardonnay Musqué wines.
Materials and Methods
Vineyard site and experimental design.
The study began in 1999 on a block of 4-year-old Chardonnay Musqué vines located at Cave Spring Vineyards in Beamsville, Ontario. Vines were grafted on SO4 rootstock and trained to a double-Guyot training system. Vines were spaced 1.2 x 2.2 m (vine x row) in rows orientated north-south. Six rows were designated for use in this trial. A randomized complete block design was used, consisting of 10-vine plots (two postlengths each) of five cluster-thinning (CT) treatments plus a nonthinned control. Thinning was performed at specific berry developmental stages—late bloom/early set, early stage I, late stage I, lag phase, and veraison—determined according to the basal cluster berry diameters of 2, 5, 7, 10, and 12 mm, respectively, combined with observations of phenological characteristics. The remaining portions of each of the rows in the field trial were left nonthinned for use in a parallel trial comprising a series of yeast strain and enzyme use treatments that could be compared to the nonthinned controls during the 1999 to 2001 seasons (Reynolds et al. 2007). No enological treatments were carried out in 2002.
Thinning treatments.
Cluster thinning for the bloom/ set, early stage I, late stage I, lag phase, and veraison treatments were performed on the respective dates of 11 and 21 June, 1 and 24 July, and 5 Aug 1999; 30 June, 14 and 27 July, and 9 and 25 Aug 2000; 27 June, 9 and 15 July, and 7 and 22 Aug 2001; and 4, 16, 22, and 29 July and 15 Aug 2002. Cluster thinning involved removing the distal cluster(s) on those shoots that produced more than one cluster per shoot, thus leaving a maximum of one cluster per shoot.
Berry sampling and harvest.
At harvest, cluster number per vine was recorded, and yield/vine was determined using a Toledo SB32000 top-loading balance (Mettler Corp., Columbus, OH). Cluster weight was determined from these data. Fruit from common treatments in adjacent blocks were subsequently combined and stored at 2°C until crushing and pressing. Approximately 15 kg of fruit was retained for winemaking from each treatment replicate.
Replicate 100-berry samples were collected each season on 15 Sept 1999, 2 Oct 2000, 17 Sept 2001, and 18 Sept 2002. These samples were later used for the determination of Brix, titratable acidity (TA), pH, and berry weight. Berries per cluster were calculated from berry weight and cluster weight data. An additional 250-berry sample was also collected for later determination of berry free volatile terpene (FVT) and potentially volatile terpene (PVT) concentration. All berry samples were stored at −24°C until analysis. Weight of cane prunings (vine size) was determined annually at pruning time by weighing the cane prunings of each vine using a dairy scale. Crop load was calculated from the ratio of yield per vine to vine size.
Winemaking.
Grapes from the CT treatments were destemmed and crushed, treated with 50 mg/L sulfur dioxide, and given 24-hr pomace contact in 20-L food-grade plastic pails at 2°C before pressing. An additional ~90 kg of fruit per row was harvested from nonthinned vines in the rows that included the thinning trial and was designated for the parallel yeast/enzyme trial (Reynolds et al. 2007). This fruit was also destemmed, crushed, and given 24-hr skin contact in each of six, 200-L stainless steel variable capacity fermentation tanks at 2°C before pressing. At the beginning of pomace contact, six 15-kg lots of crushed, destemmed grapes from nonthinned vines were set aside in 20-L food-grade plastic pails, and the enzyme product Cinn-Free (Scott Laboratories, Pickering, ON) was added to each container according to manufacturer’s recommendations. Pressing was performed in an Idropress basket/bladder press (Enoagricola Rossi, Calzolaro, Italy) at 2.0 bars pressure. Immediately after pressing, two 250-mL must samples were taken from each viticultural and enological treatment replicate for subsequent analysis of Brix, pH, TA, FVT, and PVT. Three 20-L fermentation replicates were retained for each of the six viticultural treatments and six enological treatments (three in 1999). Must from each viticultural treatment replicate was subsequently inoculated with Saccharomyces cerevisiae strain VL1 at a rate of 0.25 g/L. Enological treatments included S. bayanus strain EC1118 ± either Cinn-Free (q.v. protocol above) or AR2000 (DSM-Gist-Brocades, Servian, France) and S. cerevisiae strains CY3079, D47, and VL3. Inoculation rates were 0.25 g/L must. The CY3079, D47, and AR2000 treatments were not included in 1999, and no enological treatments were carried out in 2002. All fermentations were carried out in 20-L glass carboys at 16°C. All fermentations were allowed to proceed to 2.5 to 5.0 g/L of residual sugar, as determined by Clinitest tablets (Bayer, Etobicoke, ON). With the exception of the EC1118 + AR2000 treatment, all fermentations were arrested by addition of 75 mg/L SO2 followed by a decrease in temperature to −2°C. The designated EC1118 + AR2000 wines were cold-stabilized at −2°C, warmed to 10°C, after which AR2000 was added according to manufacturer’s directions (3 g/hL for 5 weeks). After 5 weeks, wines were relocated to a 20°C facility, and bentonite was applied to all fermentations at a rate of 5 g/hL, after which all wines were racked, cold-stabilized, and subsequently reracked. Filtration of wines was carried out using 0.5-μ pad filters (Scott Laboratories, Pickering, ON) in series with a 0.22-μ hydrophilic cartridge filter (Millipore, Bedford, MA), and immediately bottled. All wines were stored at 11°C until sensory analysis was conducted.
Berry, must, and wine composition.
Berry, must, and wine analyses were consistent with those described by Zoecklein et al. (1995). Berries and/or must samples were removed from −24°C and placed at 4°C overnight to thaw. Berries/musts were heated in an Isotemp 228 water bath (Fisher Scientific, Toronto, ON) set at 80°C for 1 hr, then allowed to cool to room temperature. Berry samples were juiced using an Omega model 500 juicer, allowed to settle, and then solids were removed through vacuum aspiration. Wine analyses were performed on bottled samples. Brix was obtained using a temperature-compensated Abbé benchtop refractometer (model 10450; American Optical Instruments, Buffalo, NY). Berry/must/wine pH was obtained using an Accumet pH/ion meter (model 25; Denver Instruments, CO). A PC Titrate (Man-Tech Associates, Guelph, ON) autotitrator was used to obtain berry/must/ wine TA using 0.1 N NaOH and a 25-mL sample. Ethanol was determined using an Agilent series 6890 gas chromatograph (Hewlett-Packard, Mississauga, ON).
FVT and PVT analysis.
Monoterpene analysis was performed based on the original method devised by Dimitriadis and Williams (1984) with modifications (Reynolds and Wardle 1989a). Berry/must samples were removed from −24°C and allowed to thaw overnight at 4°C. Wines for analysis were obtained from bottled samples that had undergone at least one year of storage at 12°C. Berry samples were homogenized in a Waring commercial laboratory blender (New Hartford, CT) for 20 sec. A 100-g berry/ must/wine aliquot was weighed in a 250-mL beaker, and pH was subsequently adjusted to ~6.7 using 20% NaOH. Samples were subsequently steam-distilled (Lurex, Vineland, NJ) and two fractions were collected: the first (25 mL) contained berry/must/wine FVT and the second (50 mL) contained the PVT. The FVT fraction was collected in 10 to 15 min, sample pH was adjusted to ~2.0 using 10 mL of 50% phosphoric acid, and the PVT fraction was subsequently collected within 15 min. The FVT fraction from the wines was set aside for further preparation before analysis. A 10-mL aliquot of each berry/must FVT and berry/ must/wine PVT distillate was transferred to a test tube and reacted with 5 mL of a 2% (w/v) vanillin solution in concentrated sulfuric acid. Test tubes were placed in a 60°C water bath for 20 min and allowed to cool at room temperature for 5 min. The absorbance of each sample was measured within 20 min at 608 nm on a Pharmacia Biotec Ultrospec UV/VIS spectrophotometer (model 1000E; Biochrom, Cambridge, UK). The monoterpene concentration of each sample was determined from a calibration curve consisting of linalool standards (0 to 10 mg/L) that had undergone the same colorimetric reaction.
Wine FVT compositional analysis.
Given the potential for interference by ethanol on wine FVT measurements (Dimitriadis and Williams 1984), the entire 25-mL FVT distillate was first diluted with 50 mL of distilled water and passed through a Sep-Pak Plus C18 cartridge (Waters Corp., Milford, MA) at no more than 5.0 p.s.i. The column was subsequently washed with 5-mL distilled water, and FVT was then eluted off the column with 100% methanol. The eluent was subsequently brought up to 25 mL with distilled water, and a 10-mL aliquot was taken for colorimetric analysis as described for the berries and musts. The calibration standards used for wine FVT analysis included 20% (v/v) of 100% methanol to discount any interference cause by this compound.
Quantitative descriptive analysis.
Judge training was conducted before formal sensory analysis, whereby subsets of wines from each component of the study were presented to judges for generation of descriptive terms. Flavor and aroma standards for each of these descriptors were then presented to the judges in subsequent training sessions and were modified until judges obtained a general consensus regarding the accuracy and intensity of the standard relative to the wine samples (Table 1⇓). Panel training was initiated annually about 6 to 8 months following harvest and about 6 weeks postbottling. Because of the large number of treatments in 1999 to 2001, a balanced incomplete block design was used for each of the formal sessions for the wines from those seasons. Each session (representing a single set of fermentation replicates) consisted of two groups of four wines (treatments) presented in random order. Each judge tasted a treatment replicate twice; once in the morning and once in the afternoon. In 2002, all six treatments were presented in each session. Twelve panelists were used each season. For details of the analysis see (Reynolds et al. 2007).
Descriptors and standards used for flavor profiling of Chardonnay Musqué wines, 1999–2002. Standards were added to 100 mL neutral white wine unless designated otherwise.
Statistical analysis.
The SAS statistical package (SAS Institute, Cary, NC) was used in analysis of yield, chemical, and sensory data. Analysis of variance was carried out using the general linear models procedure (PROC GLM). The CT treatments were additionally analyzed using single degree-of-freedom polynomial contrasts for linear, quadratic, cubic, and quartic trends. The means significantly different from the control among the CT treatments were determined by Dunnett’s t-test (Dunnett 1955). Treatment x block x vine was used as an error term for all growth and yield data, while treatment x block x postlength was used as an error term for berry samples. Interactions between viticultural and enological treatments on wine composition and sensory attributes were similarly tested by using treatment (field) x treatment (winery) x block as an estimate of error for compositional variables and treatment (field) x treatment (winery) x block x judge for sensory data. Principal component analysis (PCA) was also performed on all sensory data without rotation, as well as wine chemical data in 2000 and 2001.
Results
Impact of CT timing on yield and vine performance.
In three of four seasons (2000 to 2002), a substantial decrease in the number of clusters per vine was observed in all CT treatments based on Dunnett’s test; a significant cubic trend was also apparent for the 1999 season (Table 2⇓). Yield per vine was concomitantly reduced in four of five CT treatments in 2000, two of five in 2001, and five of five in 2002, with most notable reductions occurring when CT was performed from early stage I onward. Decreasing linear and quadratic trends were observed in yield with increased delay in CT (i.e., set to veraison) in 2000 and 2002, while no such relationships were found in 2001.
Impact of time of thinning on yield components of Chardonnay Musqué, 1999–2002.
There were small differences between the control and CT treatments for cluster weight and berries per cluster in all four seasons according to Dunnett’s test. Cluster weight tended to decrease with increased delay in timing of CT, and linear and quadratic trends were apparent in 2000 and 2001, respectively (Table 2⇑). Cluster weight data in 1999 and 2002 displayed more unusual cubic and quartic trends, respectively. Berries per cluster had decreasing linear trends with increased delay of CT in 2000 and 2002. The 1999 berries per cluster data followed a cubic trend that generally decreased as time of CT was delayed. There were no CT effects in 2001. Berry weights tended to decrease relative to increasing delay in CT. Substantial decreases in berry weights were observed when thinning was performed from early stage I to veraison in 2001.
No vine size differences were observed in 1999 or 2000, and vines did not exceed 0.5 kg cane pruning weight in either season (data not shown). In 2001, vine size increased slightly by set and early stage I treatments versus the others (1.3 kg versus 1.1 to 1.2 kg; data not shown). Crop load was noticeably lower among all CT treatments compared with the control in 2000 (4.9 to 6.0 versus 8.2), whereas in other seasons it was similar in magnitude among treatments (data not shown).
Impact of CT timing on composition.
Trends in berry soluble solids tended to be quadratic (1999 to 2001) or linear (2002) with increasing delay in CT (Table 3⇓). In 1999, early and late CT treatments increased Brix, whereas mid-season treatments decreased it relative to the control according to Dunnett’s test. In 2000 and 2002, all CT treatments were notably higher than the control in Brix, but in 2001, Brix was increased by early stage I, late stage I, and lag phase treatments only. In musts, all CT treatments were lower than the control in 1999; four, three, and five of five CT treatments exceeded the control in 2000, 2001, and 2002, respectively. A quadratic trend existed in 1999 where early and late CT treatments had highest must Brix, while in 2002, the quadratic trend seemed to favor mid-season treatments. No noteworthy trends in must Brix were found in 2000 and 2001. Despite treatment differences in berry and must Brix, ethanol concentrations in wines were not affected by treatment (Table 4⇓).
Impact of time of thinning on berry and must composition of Chardonnay Musqué, 1999–2002.
Impact of time of thinning treatments on wine composition of Chardonnay Musqué, 1999–2002.
Berry TA increased linearly with increased delay in time of CT in 1999, displayed no trends in 2000 and 2001, and displayed an unusual cubic trend in 2002 (Table 3⇑). Only one of five CT treatments differed from the control in 1999, 2000, and 2002 (either early or late stage I), and none were different in 2001. Must TA in general decreased with increased delay in CT, but followed slightly different trends than those of berries. For instance, the trend was primarily cubic in 1999 but linear in 2000 and 2001, with no apparent trend in 2002. Consequently, later CT treatments had lower must TA than the control in 1999 and 2001 (two of five treatments), while in 2000, the early stage I treatment differed from the control. Linear trends in wine TA were apparent in 2000 and 2001, but a cubic trend was evident in 2002 (Table 4⇑). In all three instances, the tendency was for wine TA to decrease relative to increased delay in time of CT. None of the treatments differed from the control in 2000, but three of five and two of five treatments were different in 1999 and 2001; one treatment (early stage I) exceeded the control in 2002 (Table 4⇑).
Berry pH showed quadratic trends in 1999 and 2001 and a primarily increasing linear trend in 2002 with respect to increasing delay in time of CT (Table 3⇑). Only one of five treatments (early stage I) had lower pH than the control in 1999, but four, two, and five of five treatments mostly exceeding the control in 2000, 2001, and 2001, respectively, according to Dunnett’s test. In musts, pH showed a predominantly decreasing quadratic trend with respect to increased delay in CT time in 1999, but a mainly increasing linear trend in 2000 and 2001 (Table 3⇑). Four of five treatments differed from the control in 1999 (all lower) and 2000 (all higher), while two of five (both stage I) exceeded the control in 2001, and all five were higher in 2002. No treatment effects were observed for wine pH in 1999, but trends in 2000 through 2002 were similar to those of the musts (Table 4⇑). In 2000, CT applied at and after early stage I resulted in a substantially elevated wine pH relative to the control, whereas in 2001, only late stage I led to an increase, while veraison CT resulted in a notable pH reduction. In 2002 all treatments exceeded the control.
Trends in berry FVT varied depending on season, but generally CT had a positive effect. A cubic trend in 1999 tended toward a decrease with increasing delay in CT; 2000 data were predominantly quadratic with highest values in early and late-thinned treatments; both 2001 and 2002 displayed increasing linear trends relative to increased delay in thinning (Figure 1A⇓). The three early treatments exceeded the control in 1999. Little to no difference existed between the control and treatments in 2000, except for lower FVT in the lag phase. In 2001 and 2002, four and five, respectively, of the five treatments exceeded the control. Trends in the must samples were somewhat different to those of the berries: 1999 data exhibited an unusual quartic trend with very little difference among CT levels; 2000 data displayed a decreasing linear trend with increased delay in thinning; 2001 showed no significant trend; and 2002 must FVT followed a quadratic (parabolic) trend with highest values in midseason treatments (Figure 1B⇓). The two midseason treatments in 1999 and 2000 were lower than the control, but late stage I timing increased must FVT in 2001, and all treatments increased must FVT in 2002. Wine FVT followed a linear trend relative to increased delay in CT in 1999, but in 2000 to 2002 showed no significant trends and exhibited little difference between any of the treatments and the control (Figure 1C⇓).
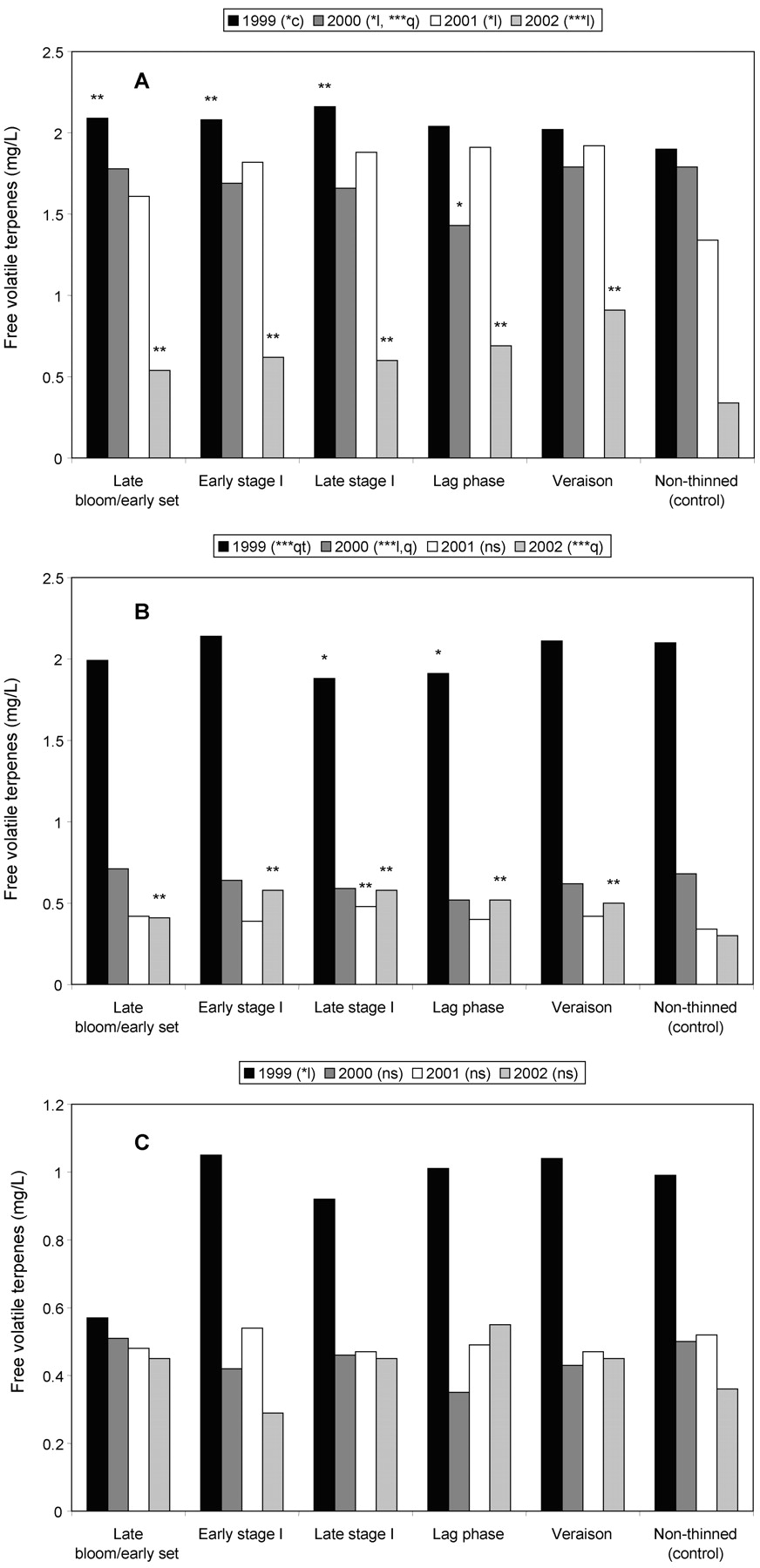
Free volatile terpene (FVT) concentrations in Chardonnay Musqué (A) berries at harvest, (B) musts, and (C) wines, as affected by time of cluster thinning, 1999–2002. l, q, c, qt: linear, quadratic, cubic, or quartic trends, respectively. ** and * indicate treatments significantly different from and less than the control, respectively (based on Dunnett’s t-test).
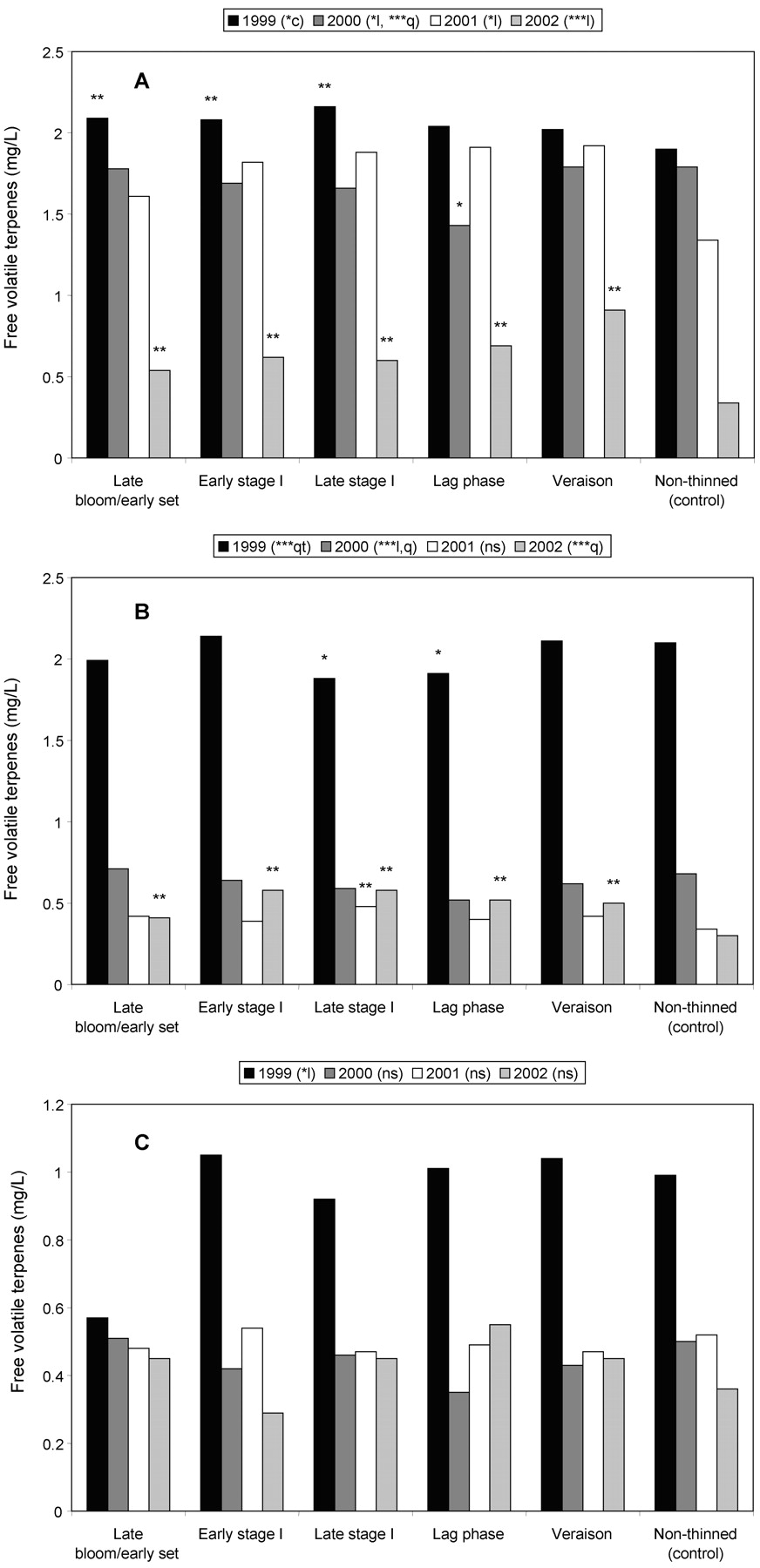
The 1999 berry PVT generally increased relative to increased delay in time of CT, and data followed a cubic trend. In 2000 to 2002, berry PVT followed either increasing linear or quadratic trends with increased delay in time of CT (Figure 2A⇓). Four of five treatments exceeded the control in 1999; early stage I and veraison CT substantially increased berry PVT over the control in 2000 (lag phase CT was lower); three midseason treatments increased PVT over the control in 2001; and all five treatments exceeded the control in 2002. Must PVT followed a cubic trend in 1999 with no easily discernible pattern (Figure 2B⇓); however, in 2000 to 2002, like the berries, it followed either increasing linear or quadratic trends relative to increased delays in time of CT. Unlike the berry results, four of five treatments in 1999 were lower than the control, but four of five (bloom/set excluded) exceeded the control in 2000, and three of five were higher in 2002. Wine PVT was not impacted by treatment in 1999 or 2002 (Figure 2C⇓). Wine PVT in 2000 and 2001 followed increasing linear and quadratic trends relative to delayed CT. Four of five treatments exceeded the control in 2000, but no differences were found in 2001.

Potentially volatile terpene (PVT) concentrations in Chardonnay Musqué (A) berries at harvest, (B) musts, and (C) wines, as affected by time of cluster thinning, 1999–2002. l, q, c, qt: linear, quadratic, cubic, or quartic trends, respectively. ** and * indicate treatments significantly different from and less than the control, respectively (based on Dunnett’s t-test).
Impact of yeast and enzyme on must and wine composition.
1999 vintage.
The addition of Cinn-Free enzyme to Chardonnay Musqué must after crushing resulted in an increase of 0.5 Brix (Table 5⇓). TA was also higher by 0.9 g/ L after addition of Cinn-Free and must pH decreased slightly. Although statistically significant, these changes may not be of practical enological significance. The addition of Cinn-Free nonetheless resulted in a greater volume of juice at pressing (data not shown). It also resulted in an increase in must FVT concentration of almost 25%, while PVT concentrations remained almost the same between treated and untreated must.
Impact of enological treatments on must and wine composition of Chardonnay Musqué, 1999–2001.
Despite a small Brix increase in Cinn-Free treated musts in 1999, ethanol concentration in the wines was not affected (Table 5⇑). Cinn-Free increased wine TA slightly concomitant with its increase in must TA, but had no effects ultimately on wine FVT or PVT. Among yeast strains, VL3 produced highest ethanol while both EC1118 treatments (± Cinn-Free) were lowest. Wines produced with VL1 had slightly lower wine TA than other treatments and retained highest PVT, but pH and FVT were not affected by yeast strain.
2000 and 2001 vintages.
Cinn-Free reduced must soluble solids by 0.2 Brix in 2000 but had no impact on must TA or pH (Table 5⇑). Inexplicably, both must FVT and PVT were reduced by addition of Cinn-Free in 2000. No significant effects of Cinn-Free on must composition were observed in 2001 (data not shown). Cinn-Free did not increase ethanol concentration in wines compared with EC1118 alone, but wines produced using EC1118 ± Cinn-Free had higher ethanol than AR2000 wines in 2000 and 2001. Cinn-Free consistently increased wine TA relative to EC1118 alone in both years, as it did in 1999. It led to increased and decreased wine pH in 2000 and 2001, respectively, relative to EC1118 alone. As with 1999 musts, substantial increases in wine FVT were observed in Cinn-Free-treated wines relative to EC1118 alone in both years. Although no differences were observed in 2000, wines to which Cinn-Free had been added (to musts) exhibited substantially lower PVT concentrations in 2001. Addition of the enzyme AR2000 to EC1118 wines postfermentation increased wine TA in 2000 and consistently decreased pH in both vintages. AR2000 decreased wine FVT in 2000 but had little impact on wine FVT in 2001. However, EC1118 fermentations to which AR2000 was added exhibited substantially lower PVT concentrations in 2001.
Wines fermented with EC1118 ± Cinn-Free or AR2000 had highest ethanol in 2000 and 2001, while VL1 and VL3 wines were lowest in 2000; VL3 wines also had lowest ethanol in 2001 (Table 5⇑). Wines fermented with VL1 had highest TA in 2000 and 2001. Across the two vintages studied, wines produced using D47 consistently displayed the lowest pH values while VL1 displayed the highest. In both years, yeast strain failed to have a substantial impact upon wine FVT. Greater differences were observed in wine PVT as a result of yeast strain selection, with VL1-fermented wines exhibiting the lowest concentrations in both years, and EC1118 generally exhibiting the highest.
Impact of treatments on sensory properties.
1999 season.
There were differences in sensory properties among CT treatments in 1999 (Table 6⇓). Dried fruit aroma was decreased by all CT treatments relative to the control, late stage I CT decreased citrus aroma, and grassy aroma was increased by bloom and veraison CT. Little difference between CT treatments and the control were observed in 1999 for all flavor and mouthfeel attributes, although color was reduced by all CT treatments relative to the control.
Impact of time of thinning treatments on sensory attributes of 1999 Chardonnay Musqué wines.
With the exception of lychee aroma, all sensory attributes from the 1999 vintage were affected by enological treatments (Table 7⇓). Wines fermented with the VL3 were highest in spicy aroma, floral, spicy, and grassy flavors, and had highest astringency. EC1118 was lowest in body and color but equal to the other yeast strains in most other attributes. Addition of Cinn-Free resulted in wines perceived as lowest in floral, citrus, and grassy aromas, citrus and grassy flavors, and astringency, but with highest dried fruit aroma, lychee and dried fruit flavors, body, finish, and color.
Impact of enological treatments on sensory attributes of 1999 Chardonnay Musqué wines.
The first two factors of PCA of the 1999 vintage explained 74.2% of the variation exhibited by the wines (Figure 3⇓). Factor I was explained primarily by grassy and citrus aromas and flavors, astringency, and acidity, which were inversely correlated with dried fruit and lychee aroma and flavor attributes, color, body and length of finish. Floral and spicy aromas and flavors and body were aligned along factor II and correlated with each other. All CT treatments were located to the left of factor II and below factor I, where they were characterized by lack of spicy, dried fruit, and lychee aromas, spicy and floral flavors, and lack of length of finish and depth of color. Generally, little separation of the viticultural treatments existed. There was greater variability among the wines according to enological treatment. VL3 was located high along factor II and to the left of factor I and was thus described primarily by floral aroma and spicy aromas and flavors. EC1118 + Cinn-Free was located to the right of factor II and below factor I and was characterized by high dried fruit and lychee flavors. VL1 (the nonthinned viticultural control) and EC1118 were grouped to the left of factor II and below factor I, where they were described by similar attributes to the CT treatments.

Principal component analysis of Chardonnay Musqué wines from nine treatments, comprising six time of thinning and three enological treatments, 1999. The VL1/control treatment also represents the nonthinned viticultural treatment. Lowercase variables are aroma attributes and uppercase variables are retronasal, taste, and color attributes.
2000 season.
In the 2000 vintage, CT timing resulted in few substantial differences among wines, mainly in terms of dried fruit aroma, muscat flavor, and color, all of which increased with increased delays in CT (Table 8⇓). Non-thinned vines, however, differed substantially from CT vines only in terms of color, whereby all CT treatments exhibited greater color intensity.
Impact of time of thinning treatments on sensory attributes of 2000 Chardonnay Musqué wines.
Tropical fruit aroma, muscat flavor, and color varied as a result of yeast strain selection and enzyme use (Table 9⇓). Tropical fruit aroma was highest among wines produced by CY3079 and EC1118 + AR2000 treatments and lowest in wines produced with VL1. Muscat flavor was highest in EC1118 + AR2000 wines and lowest in D47 wines. VL1 wines had highest color, while CY3079 had lowest.
Impact of enological treatments on sensory attributes of 2000 Chardonnay Musqué wines.
The first two factors of PCA explained 47.9% of the variation between wines (Figure 4⇓). Factor I was primarily explained by muscat aroma, floral, citrus, tropical fruit and tree fruit aromas and flavors, and sweetness, as well as wine FVT, which were inversely correlated with honey aroma, muscat flavor, astringency and color sensory attributes along with wine pH. Dried fruit and acidity and wine PVT were aligned along factor II and were inversely correlated with grassy aroma, body, and wine TA. A greater difference between CT treatments was observed in 2000 compared with 1999. Wines from nonthinned and early CT treatments were associated with greater grassy aroma and body, whereas wines from later CT treatments were associated with honey aroma, muscat flavor, astringency, and color. As in the 1999 vintage, wines from the 2000 growing season were highly variable in character relative to enological treatment. Both EC1118 and all VL1 fermentations were located to the left of factor II and were thus separated from the remaining enological treatments. These treatments were described primarily by honey and grassy aromas, muscat flavor, astringency, body, and color intensity. Wines from D47, VL3, and EC1118 + Cinn-Free treatments were located to the right of factor II and below factor I, where they were described by muscat, citrus, and tree fruit flavors and high FVT and TA. CY3079 and EC1118 + AR2000 treatments were also located on the right of factor II but above factor I and were associated primarily with dried fruit, tropical fruit, floral, and muscat aromas, tree fruit and tropical fruit flavors, sweetness, acidity and high PVT.

Principal component analysis of Chardonnay Musqué wines from 12 treatments, comprising six time of thinning and six enological treatments, 2000. The VL1/control treatment also represents the nonthinned viticultural treatment. Lowercase variables are aroma attributes and uppercase variables are retronasal, taste, and color attributes.
2001 vintage.
Several attributes were substantially affected by viticultural treatments in 2001 (Table 10⇓). Delayed CT tended to increase citrus and apple/pear aromas and decrease vegetal aroma, but flavor attributes showed no discernible pattern. Comparison with nonthinned vines, set and late stage I CT substantially reduced apple/pear aroma, while veraison CT notably reduced vegetal aroma and peach flavor, but increased herbaceous/grassy flavor. Veraison CT decreased sweetness, body, and color and increased acidity relative to the control. Both sweetness and color intensity were also substantially lower in wines produced by all but late stage I CT vines, while body and acidity were higher than the nonthinned controls in the lag phase and early stage I treatments, respectively.
Impact of time of thinning treatments on sensory attributes of 2001 Chardonnay Musqué wines.
In enological treatments, wines from the VL1 treatments were highest in vegetal, herbaceous/grassy, and mineral aromas and lowest in floral and muscat aromas, body, and sweetness (Table 11⇓). Both VL1 and VL3 produced wines lowest in peach aroma and acidity. EC1118 + AR2000 was low in peach and herbaceous/grassy aromas. Vegetal aroma was lowest among wines in EC1118 and EC1118 + Cinn-Free treatments. Sweetness was also low in EC1118, EC1118 + Cinn-Free, and EC1118 + AR2000 treatments. Wine color was of highest intensity in VL1 and EC1118 treatments and lowest in the CY3079 treatment.
Impact of enological treatments on sensory attributes of 2001 Chardonnay Musqué wines.
The first two factors of PCA explained 66.3% of the variation exhibited by the wines from the 2001 vintage (Figure 5⇓). Factor II was explained by apple/pear, muscat, and peach flavors and color, which were inversely correlated with grassy/herbaceous flavor. Factor I was explained by peach, apple/pear, citrus, muscat, and floral aromas, citrus flavor, and acidity, which were inversely correlated with vegetal, mineral, and herbaceous aromas and vegetal flavor, body, and sweetness. Delineation among viticultural treatments was observable along factor I as well as along factor II. Wines produced from non-thinned vines were located highest along and to the left of factor II and were generally differentiated from wines produced from the other CT treatments, which were associated primarily with vegetal, mineral, and herbaceous aromas and vegetal flavor, and hence low in apple/pear, citrus, muscat, and floral aromas. The veraison treatment was of particular interest, being located the closest to factor II and furthest below factor I, and was hence characterized by high herbaceous/grassy flavor, low peach, apple/pear, and muscat flavors, and low color.

Principal component analysis of Chardonnay Musqué wines from 12 treatments, comprising six times of thinning and six enological treatments, 2001. The VL1/control treatment also represents the non-thinned viticultural treatment. Lowercase variables are aroma attributes and uppercase variables are retronasal, taste, and color attributes.
Wines in 2001 exhibited large variation according to enological treatment (Figure 5⇑). Those produced using VL1 were located to the left of factor II, where they were generally characterized by a greater intensity of mineral, herbaceous/grassy, and vegetal aromas, vegetal flavor, body, and sweetness. The remaining wines were located to the right of factor II, with EC1118, EC1118 + Cinn-Free, and EC1118 + AR2000 located above factor I, and VL3, CY3079, and D47 located below factor I. EC1118 wines with and without addition of enzymes therefore tended to be associated with muscat, apple/pear, and citrus flavors, whereas VL3, CY3079, and D47 wines tended to be higher in grassy/herbaceous flavor.
2002 season.
In general, treatments that were not thinned or CT early in the growing season produced wines characterized by high citrus flavor, high acidity, and low overall impression on judges (Table 12⇓). Wines made from control vines were also characterized by low earthy/mushroom aromas and flavors. Vines thinned at berry set produced wines with a higher citrus flavor and acidity than any other wines from the thinning trial and also had low earthy/mushroom aroma and flavor. Wines produced from vines CT midseason displayed highest earthy/mushroom aromas and flavors. Wines made from early stage I CT had the lowest overall impression on judges and were characterized by low intensities in all variables, except for earthy/mushroom aromas and flavors, and high acidity. Wines from late stage I CT similarly had low intensities for most variables other than earthy/mushroom aroma and flavor, and a low overall impression on judges. Wines from treatments thinned late in the season had highest overall impression on the judges. Wines from lag phase CT had high earthy/mushroom flavors and aromas, low citrus flavor, and a lower overall impression on tasters. Wines from vines thinned at veraison had low earthy/mushroom aroma and flavor, a high intensity of perceived acidity, and highest overall impression on judges.
Impact of time of thinning treatments on sensory attributes of 2002 Chardonnay Musqué wines.
Discussion
Impact of CT timing on yield and vine performance.
Thinning substantially reduced the number of clusters per vine and was undoubtedly the main contributing factor to reductions in yield and crop load. Yield was not as substantially reduced in the bloom/set treatment compared to later treatments. Given that assimilate demand by clusters increases rapidly following berry set, CT at or before this phenological stage has resulted in a compensation response in some yield components (Edson et al. 1995, Miller et al. 1996, Reynolds 1989, Reynolds et al. 1994a). In one study, vines thinned at 2, 4, and 6 weeks after set had lower berry weights than vines thinned at and before berry set (Weaver and Pool 1973). In another study, thinning before set increased the number of berries per cluster compared with vines thinned subsequent to set (Smithyman et al. 1998). The primarily decreasing linear relationships between increased delay of thinning time and yield, cluster weight, berries per cluster, and berry weights suggest a similar compensation response in this study for vines thinned early in the season.
Competition also exists between reproductive and vegetative growth, whereby vegetative growth is sacrificed at the expensive of reproductive growth (Edson et al. 1995). In this case, the slight increase in vine size among early CT treatments in relation to the control was expected.
Effect of CT timing on composition.
In most cases berry and must Brix was increased over the nonthinned controls and Brix tended to increase with increased delay in thinning time. The relationships were mainly quadratic in 1999 to 2001, but linear (berries only) in 2002. Similar trends in soluble solids relative to time of thinning have been observed in Thompson Seedless (Weaver and Pool 1973) and Chardonnay (van Schalkwyck et al. 1995). The responses of berry and must TA were not large and might be attributed to accelerated berry maturity in the CT treatments. Data suggested a general trend toward decreased TA with later application of CT in the season. These results tend to agree with observations that TA is affected very little by thinning (Edson et al. 1995, Reynolds et al. 1994a, Smithyman et al. 1998, Weaver and Pool 1973). In three of four seasons, some thinning treatments increased berry, must, and wine pH when performed at and subsequent to early stage I. The increased pH observed in some of the CT treatments after set are similar to results obtained on Seyval grapevines (Smithyman et al. 1998). The increase in pH during the 2000 and 2002 seasons with CT at and subsequent to early stage I may have been due to increased potassium flux into a lesser number of sinks (Hepner and Bravdo 1985), thus increasing pH.
Berry and must FVT responded to CT as well; in three of four seasons, there was either an increase relative to increased delay in time of thinning or an increase in FVT in CT treatments over the control. In general, FVT have tended to be unresponsive to viticultural treatments (McCarthy 1986, Reynolds and Wardle 1989a,b), with little variation observed in Riesling FVT relative to crop level (Reynolds et al. 1994a). Given that no differences in wine FVT were observed between treatments from 2000 to 2002, it appears that any differences that may have been brought about by CT in berry and must FVT were of little consequence to final wine FVT concentration.
Berry, must, and wine PVT were all increased by CT in some seasons, and an increase in PVT with increasing delay in thinning time was observed. Unlike FVT, wine PVT was positively impacted by CT in two of four seasons. PVT has been very responsive to canopy management and thinning practices (McCarthy 1986, Reynolds and Wardle 1989a). Increased PVT with reduction in crop level has generally been ascribed to advancements in berry maturity, which this study tends to confirm, since berry Brix and pH values in most CT treatments were also concomitantly higher than nonthinned treatments. PVT concentration therefore may be similarly responsive to changes in crop level as observed in Brix and pH.
Impact of yeast and enzyme on wine composition.
Strain VL1 tended to exhibit increased TA over the other yeast strains in 2 of 3 years, perhaps because of variation among yeast strains to metabolize organic acids such as malic, succinic, citric, and lactic acids (Antonelli et al. 1999, Reynolds et al. 2001). Provided that TA often correlates inversely to pH, it is interesting that wines produced from VL1 generally exhibited increased pH values over the other yeast strains in 2 of 3 years, which may have been due to variation among wines in concentrations of individual organic acids, particularly malic acid.
Yeast strains have exhibited the potential to alter terpene concentration, and thus wine aroma, through biosynthesis of terpenes, biotransformation of free terpenes, and/or enzymatic hydrolysis of terpene glycosides. For instance, Saccharomyces fermentati, a yeast species used in the production of fino sherry, produced trace amounts of linalool, cis-neroldiol, trans-neroldiol, and trans, trans-farnesol when growing as surface film on a simulated wine medium. The close relatedness of this species to S. cerevisiae led the authors to speculate that S. cerevisiae may also have such ability (Fagan et al. 1981). This assumption was further supported in a study in which geraniol, linalool, α-terpineol, and (E)- and (Z)-neroldiol accumulated within the yeast cell during fermentation with a S. cerevisiae strain and two film-forming yeasts (S. bayanus, and S. cerevisiae var. capensis) (Zea et al. 1995). The ability for S. cerevisiae to convert linalool, nerol, and geraniol into α-terpineol in synthetic medium was also noted (King and Dickinson 2000).
A number of different yeast strains have displayed β-glycosidase activity. Various degrees of β-glucosidase activity were observed in several non-Saccharomyces species (McMahon et al. 1999), while other studies have indicated that Saccharomyces exhibit little to no β-d-glucosidase activity (Rosi et. al. 1994). For example, 74 strains of Saccharomyces were screened, and although esterase, protease, and pectinase activities were found, no β-glucosidase activity was observed (Ubeda Iranzo et al. 1998). However, McMahon et al. (1999) observed β-glycosidase activity in S. cerevisiae VL1 strain. Similarly Delcroix et al. (1994) observed α-arabinosidase, α-rhamnosidase, and β-glucosidase activity in three Saccharomyces strains, the activity of which was most evident during the exponential growth phase of fermentation. Mateo and Di Stefano (1997) found varying degrees of activity among six Saccharomyces strains. Grossmann and Rapp (1988) found that yeast β-glucosidase activity could be maintained at 90% of its optimal activity 3 weeks after completion of fermentation; however, the majority of research suggests that yeast β-glucosidase activity is limited because of pH ranges, ethanol concentrations, and temperatures most commonly present during the production of wine (Delcroix et al. 1994, Mateo and Di Stefano 1997, Rosi et al. 1994).
Wine FVT in this study, despite previous evidence otherwise, was influenced very little by yeast strain selection. Lack of substantial increase in FVT in wines made from yeast strains exhibiting correspondingly low PVT, such as VL1, which purportedly has substantial glycosidase activity, also suggests hydrolysis of wine PVT does not necessarily result in a concomitant increase in FVT. Zoecklein et al. (1997) suggested that absorption, isomerization, and metabolism of terpenes by yeast strains may contribute to the lack of observed increase in specific free monoterpenes with reduction in their glycosidic counterparts.
Enzyme addition, particularly Cinn-Free, resulted in substantial increases in must and wine FVT. It appears that addition of this pectinase enzyme preparation to crushed grapes before pressing will potentially increase wine FVT, presumably through either greater extraction of FVT from the skins where the bulk of secondary metabolites are located (Wilson et al. 1986) or β-glucosidase side activity. The increase in wine TA by this treatment may similarly be the result of greater extraction of organic acids located within the skin of the berry. Although both AR2000 and Cinn-Free decreased wine PVT, the lack of increase in wine FVT by AR2000 suggests that the β-glucosidase side activity by which this product is marketed was insufficient for increasing wine FVT.
Impact of treatments on wine sensory attributes.
The sensory profile of wine is dependent on aroma and flavor compounds produced in the grape, during fermentation, and through aging (Rapp and Mandery 1986). Several studies using thinning have found reasonable correlations between terpene concentration in grapes or wines (McCarthy 1986, Reynolds et al. 1994a,b) and wine sensory response. Ethanol, higher alcohols, and acetate and ethyl esters are among the most common fermentation-derived compounds that contribute to wine aroma (Dumont and Dulau 1996, Rapp and Mandery 1986). Formation of these compounds has been reported to be highly dependent on yeast strain. In one study, S. bayanus strains produced substantially higher volatile content than S. cerevisiae, mostly because of higher concentrations of phenylethanol and ethyl lactate (Antonelli et al. 1999). Differences in Riesling and Chenin blanc wine aromas were also reported as a result of several strains of S. cerevisiae and S. bayanus (Reynolds et al. 2001). Given such variation in wine chemical composition with yeast strain selection, reports of an influential effect of yeast strain selection on wine sensory profiles are not surprising (Dumont and Dulau 1996, Reynolds et al. 2001).
Despite the physiological constraints placed on β-glucosidases by the wine medium, wine chemical and sensory properties may still be impacted by use of different commercial enzymes. Through addition of Novoferm 12, an Aspergillus niger pectinase preparation containing α-rhamnosidase, α-arabinosidase, and β-glucosidase activity, Canal-Llaubères (1994) observed increased free monoterpenes in both Gewürztraminer and muscat wines. Similar increases to free monoterpenes in Morio-Muskat wines with addition of commercial enzymes were also reported by Grossmann and Rapp (1988), who found that the enzyme-treated wines had higher varietal typicity and received higher sensory scores compared with non-enzyme-treated wines. Impact of enzyme addition on wine chemical composition and sensory attributes appears inconsistent, however. For instance, through the addition of Novoferm 12, Hesford et al. (1995) failed to find increases in free monoterpenes of Müller-Thurgau wines similar to those of Canal-Llaubères (1994). In Semillon wines produced with and without the pectolytic enzyme preparation Rohapect C, only subtle sensorial differences were found, despite a reduction in glycosyl-glucose concentration from the juice to the wine (Francis et al. 1996).
In this study—with the exception of dried fruit, spicy, citrus, and grassy aromas, citrus and spicy flavors, and color—thinning or its timing had little effect on wine sensory properties for the 1999 vintage. However, yeast strain selection and enzyme addition strongly impacted all sensory attributes except lychee aroma. Individual yeast strains had very specific impacts upon aroma and flavor; for instance, wines produced using VL3 were generally highest in spicy, grassy, and floral aromas, spicy flavor, astringency, and body than those produced using either EC1118 or VL1. All VL1 wines were associated mainly with grassy, citrus, and astringency descriptors. Little differences in sensory attributes were observed between EC1118 alone and VL1. Addition of Cinn-Free before pressing, however, increased dried fruit aroma, lychee, dried fruit flavor, and length of finish, and decreased grassy aroma, citrus, and grassy flavors compared with wines produced solely with EC1118.
As in 1999, CT had little effect on wine attributes in 2000. PCA suggested that grassy aroma and body were associated mainly with wines from nonthinned vines and those thinned at bloom/set. A slightly greater effect upon sensory attributes was exerted by yeast strain and enzyme use. Among the enological treatments, VL1 wines were considerably lower in tropical fruit aroma, and were again associated with grassy aroma and astringency.
In 2001, the veraison CT treatment was noteworthy given its reductions in vegetal aroma and peach flavors and increased intensity of herbaceous/grassy flavor. Both the control and late stage I wines exhibited greater depth of color and a higher perception of sweetness than the other treatments. Increased acidity in wines produced from lag phase and veraison-thinned vines and decreased body in those produced from early stage I and veraison-thinned vines further differentiated the treatments. Yeast strain and/or enzyme use again had a greater magnitude of effect. Wines produced using VL1 were again clearly separated from the remaining enological treatments. Wines produced with VL1, along with those from VL3, were also perceived as unique from the other enological treatments.
In 2002, again very few sensory attributes were impacted by time of thinning, with the exception of overall impression, which was highest in veraison-thinned wines. Sensory data from all four years indicated that the sensory characteristics of these wines were more profoundly affected by enological treatments than by cluster thinning, which is noteworthy because large decreases in yield (and increased labor costs) can result from cluster thinning. It is also significant that many differences in berry and must Brix, TA, pH, and even FVT and PVT created by CT had relatively small impacts on the perception of sensory attributes. Very few aroma and flavor attributes (floral and muscat excepted) that distinguished one viticultural treatment from another were highly correlated with either FVT or PVT. That is not surprising with FVT, since little differences existed in berries, musts, and wines among CT treatments and between CT treatments and the control.
Enological treatments were distinguishable in tropical fruit, floral, and peach aroma attributes, which were also correlated with wine PVT. VL1 wines were particularly noteworthy by being lower in intensity in all of these attributes than the other enological treatments. Although PVT are odorless, and it is therefore doubtful that increased PVT alone increased the floral, fruity, and peach aromatics of these wines, it may be that hydrolysis subsequent to their analysis may have impacted sensory perception. It is also equally possible that some yeast strains were capable of permitting isomerization among various monoterpene moieties. For example, Zoecklein et al. (1997) observed that α-terpineol increase, and geraniol and nerol decreased subsequent to fermentation, the degree of which was somewhat dependent on the yeast strain used. The lower PVT observed among VL1 fermentations in two of three seasons may therefore indicate that VL1 has the capability not only to convert PVT to their free forms but also to biotransform free terpenes. Greater conversion of several monoterpenes into α-terpineol and citronellol, for instance, may have lessened fruity and floral aromas and enhanced grassy, vegetal, and citrus attributes. Further research is needed to support such a claim.
Conclusions
Cluster thinning of Chardonnay Musqué grapevines in Ontario led to reductions in yield, enhancements in berry, must, and wine composition, yet very modest changes in wine sensory attributes. Such results tend to indicate that thinning had relatively little impact on final wine aroma and flavor despite effects on berry, must, and wine composition. It was instead apparent that yeast strain selection and use of enzyme products have the potential to create profound differences in wine sensory properties. The correlation of chemical variables with some sensory attributes tends to imply that alteration in chemical composition by yeast strains and/or enzyme products may have caused some of the perceived differences among treatments. Differences obtained in the vineyard through cluster thinning tended to indicate advanced berry maturity. Such advancement may be of little benefit to growers of midseason cultivars such as Chardonnay Musqué, however, as it is rarely difficult to achieve high soluble solids with these aromatic cultivars. Results suggest that costs associated with application of these viticultural treatments in terms of labor and losses in yield are of little benefit to grapegrowers and winemakers.
Footnotes
Acknowledgments: Financial assistance from the Natural Sciences and Engineering Research Council and the National Research Council of Canada is hereby acknowledged.
The authors thank Angelo Pavan and Kevin Latter, Cave Spring Estate Winery, for their cooperation. Efforts of the many sensory panellists are also acknowledged. This paper represents work included in the MSc thesis of J. Schlosser (2003) and the undergraduate theses of D. Sorokowsky (2000) and R. Roberts (2003).
- Received May 2005.
- Revision received October 2005.
- Revision received July 2006.
- Copyright © 2007 by the American Society for Enology and Viticulture
Literature Cited
Vol 58 Issue 1








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
More from this TOC section
Similar Articles
