


Abstract
Five certified clones of Merlot from Foundation Plant Services (FPS), University of California, Davis, were evaluated for viticultural performance in Oakville, Napa Valley. Vine yield components (cluster number, cluster weight, clusters per shoot, berries per cluster, and berry weight), fruit composition (Brix, pH, titratable acidity), potassium (K) concentration, and vegetative growth parameters (pruning weight and shoot weight) were measured over five growing seasons, 1997 to 2001. Yield, berries per cluster, berry weight, and cluster weight varied significantly among clones, and all yield components varied significantly from season to season. FPS 8 yielded 35% less than clones FPS 1, 3, and 6. FPS 9 showed the most erratic yield pattern over seasons. The number of berries per cluster was primarily responsible for yield differences. Fruit composition parameters pH and K concentration varied significantly among clones, with FPS 8 having both the highest pH and K concentration. Brix concentration at harvest did not significantly differ among clones despite yield differences. Vegetative growth differed significantly among clone, with FPS 8 having the greatest pruning weight and FPS 1 the least. Yield to pruning weight ratios were lowest for FPS 8 and highest for FPS 1.
Merlot is one of the most important winegrapes in California and is currently the third most widely planted wine-grape with 21,660 ha (53,530 acres) (CASS 2005). Second only to Cabernet Sauvignon in planted acreage of red varietals, Merlot’s status in California viticulture makes it an important research subject.
A growing number of researchers have reported on the type and amount of variation among clonal selections of winegrape varieties. Studies have shown clonal differences in yield, cluster architecture, vegetative growth, fruit composition and wine composition (Wolpert 1996, Wolpert et al. 1994, 1995, Cirami and Ewart 1995, Kotseridis et al. 1998, Bettiga 2003a,b). Historically in California, clones selected and released by Foundation Plant Services (FPS), University of California, Davis (UC Davis) were not accompanied by information on their viticultural characteristics (Wolpert 1995). For this reason, studies have been underway to evaluate FPS clones of important varieties such as Chardonnay, Pinot noir, Cabernet Sauvignon, and Zinfandel (Wolpert 1995).
Clonal evaluations of Merlot have been ongoing in Bordeaux, France, for over 30 years (Delmas 1995, Leclair 1995). These studies have focused on selecting clones for superior wine quality while maintaining vine health and performance. Naturally, this work has focused on French selections and thus is not directly relevant to data on selections that do not have a direct link to France.
The purpose of this trial was to evaluate viticultural characteristics of five FPS Merlot clones in Oakville, an American Viticultural Area (AVA) in Napa Valley, California. Oakville is widely regarded as one of the top AVAs in California for the production of quality winegrapes, particularly red Bordeaux varieties. Four of the five clones were previously evaluated at other sites in California (Roberts and Blazer 1995, Bettiga 2003a). Ideally, clones should be evaluated in multiple sites because clonal characteristics such as yield and maturity can vary significantly from site to site (Cirami and Ewart 1995).
Materials and Methods
Five Merlot clones, FPS 1, 3, 6, 8, and 9, as listed in Table 1⇓, were evaluated for viticultural performance in this study. Vines were planted in 1991 at the Oakville Station Vineyard in Oakville, California, operated by the Department of Viticulture and Enology, UC Davis. The soil was a deep, gravelly Bale clay loam with less than 2% slope (Lambert and Kashiwagi 1978). Vines were grafted onto Teleki 5C rootstock (Vitis riparia x V. berlandieri) and were planted to a 1.8 m x 2.4 m spacing (6 ft x 8 ft), equivalent to a vine density of 2,242 vines per ha (907 vines per acre). Vines were trained to a bilateral-cordon vertical shoot-position (VSP) system with a single fruiting wire at 100 cm aboveground and paired dual shoot-positioning wires at 145 and 190 cm aboveground. A drip system was used for supplemental irrigation. Other cultural practices were consistent with Napa Valley winegrape vineyards.
Source and heat treatment of Merlot clones from Foundation Plant Services (FPS), planted in Oakville, Napa Valley, CA.
Data collection began in 1995 (Barnhisel 1998) when the vines were five years old and continued through 2001. However, vines in 1995 and 1996 were not fully mature, as determined from shoot number per vine (data not shown) and only data from 1997 to 2001 were used. In all years, harvest occurred on a single day, although the date varied from year to year. Vines were harvested individually, clusters were counted, and vine crop weights measured. Prior to harvest, one 100-berry sample was taken per treatment replicate. Samples were returned to the laboratory where they were weighed to determine berry weight. Berries were crushed and juice was filtered through cheesecloth. Soluble solids were measured as Brix with a hand-held, temperature-compensating refractometer. Juice pH was measured with an electronic pH meter, and juice samples were titrated with 0.1 N NaOH to pH 8.2 endpoint to determine titratable acidity (TA). Juice potassium (K) concentration, as determined by atomic absorption analysis, was measured in all years except 2001.
During the dormant season, shoots on each vine were counted. The weight of prunings was measured and used to calculate the average weight per shoot and the yield: pruning ratio. Vine shoot number varied from 22 to 26 shoots per vine depending on year, but was kept consistent among clones in any given year.
A randomized complete block design was employed, with eight replicates of 27 vines each, arranged as experimental units of three rows by nine vines. The centermost five vines of each replicate were used for data collection. For statistical purposes, the experiment was treated as a split-plot design where clones were the main plot and years were subplots. Data were analyzed by ANOVA with mean separation by Duncan’s multiple range test.
Results
Yield components.
Yield was significantly lowest for FPS 8, which averaged 5.6 kg per vine as compared to FPS 1 (8.1 kg), FPS 3 (8.4 kg), FPS 6 (8.5 kg), and FPS 9 (7.8 kg) (Table 2⇓). Yields varied significantly from year to year as did all yield components (Table 2⇓). Seasonal variation affected clonal yields differently, as indicated by the highly significant interaction of clone by year. FPS 9, second lowest in average yield, had the most erratic yield pattern (Figure 1A⇓), with the highest and second highest yield in 1997 and 2000, respectively, and the second lowest yield in 1998 and 2001.
Yield components of five Merlot clones planted in Oakville, CA. Clonal main effects are averages of five years (1997 to 2001) and year main effects are averages of the five clones.

Yield per vine (A) and berries per cluster (B) of five Merlot clones planted in Oakville, CA, 1997 to 2001.
Shoot number per vine was imposed by pruning and shoot-thinning practices to the same level for all clones and was not related to differences in yield (Table 2⇑). Clusters per shoot and clusters per vine were nearly identical among clones and were unrelated to yield differences.
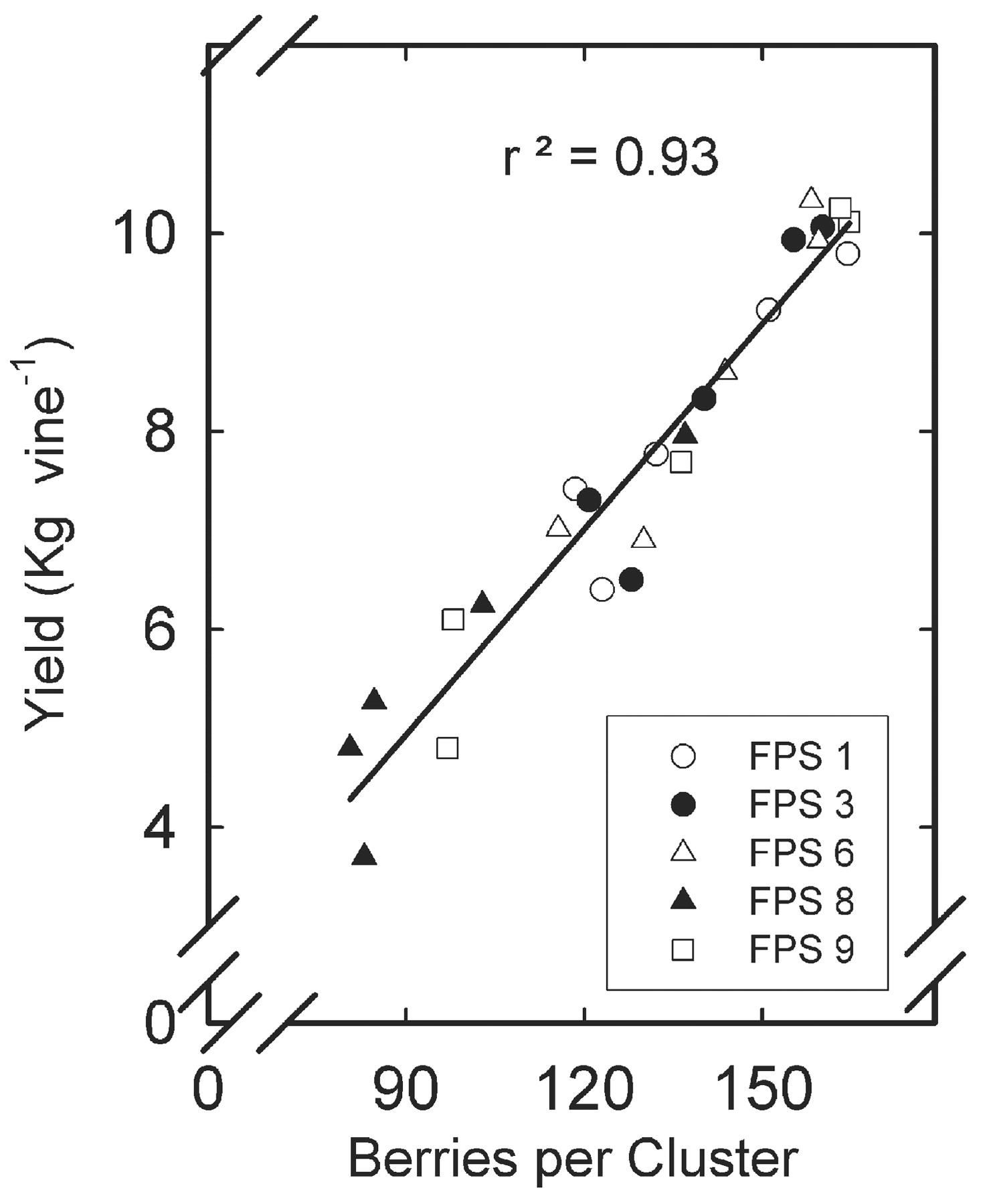
Cluster weight differed among clones by more than 50%, from a low of 138 g for FPS 8 to a high of just over 200 for FPS 1, 3, and 6 (Table 2⇑). Cluster weight differences were primarily due to numbers of berries per cluster and, to a much lesser degree, berry weight. The number of berries per cluster varied from a low of 98 for FPS 8 to a high of 141 for FPS 3 and 6 (Table 2⇑). A nearly equivalent range of values was seen among the five growing seasons, with berries per cluster ranging from a low of 108 in 1998 to a high of 157 in 1997. Mirroring its erratic pattern in seasonal yield, FPS 9 was the most inconsistent in berries per cluster, being second lowest 1998 and 2001 and in highest in 1997 and 2000 (Figure 1B⇑). Number of berries per cluster was highly correlated with yield (r2 = 0.93) for all clones (Figure 2⇓). Berry weight played a small but significant role in clonal yield, with average differences being 0.1 g (Table 2⇑). However, differences in berry weight were greater among years with 0.4 g variation from lowest at 1.3 g in 1999 and 2001 to highest at 1.7 g in 1997. However, the lack of a significant interaction term (clone*year) for berry weight indicates that, unlike berries per cluster and cluster weight, the differences were consistent among clones from year to year.

Correlation between berries per cluster and vine yield for five Merlot clones planted in Oakville, CA. Data points are individual years (1997 through 2001).
Fruit composition.
Fruit chemistry parameters Brix and TA were not significantly different among clones (Table 3⇓). In any one year, there was no more than a 0.5 degree Brix difference among clones. FPS 8 had higher juice K concentration and higher juice pH at harvest. Year-to-year differences in fruit composition parameters were due to timing of harvest.
Fruit composition of five Merlot clones planted in Oakville, CA. Clone main effects are averages of five years (1997 to 2001), except for potassium (K), which is a four-year average (1997 to 2000). Year main effects are averages of the five clones.
Vegetative growth.
FPS 8 produced the greatest vegetative growth, as measured by pruning weight, 1.9 kg per vine (Table 4⇓). FPS 1 had lowest pruning weight at 1.5 kg, followed by FPS 9 at 1.6 kg. Because shoot number per vine was regulated as a cultural practice, shoot weight directly reflected pruning weight. FPS 8 had the greatest shoot weight at 84 g, while FPS 1 and 9 had the lowest shoot weights at 66 g and 69 g, respectively. Because of large variation, differences among years were not significantly different for both pruning weight and shoot weight. FPS 8 had the lowest yield-to-pruning-weight ratio, 3.1. That was a function of both its lower yields and higher pruning weights. FPS 1, 3, 6, and 9 all had yield-to-pruning weight ratios between 5 and 6.
Vegetative growth components for five Merlot clones planted in Oakville, CA. Clone main effects are averages of five years (1997 to 2001) and year main effects are averages of the five clones.
Discussion
The data presented here on Merlot clone performance is the third such study in California. Bettiga (2003a) compared four Merlot clones in Monterey County, on California’s Central Coast, an area cooler and windier than Oakville. In the Oak Knoll locale in Napa Valley, Roberts and Blazer (1995) compared four certified clones to two field selections preferred by their winery. While their site bears some similarity to the Oakville site presented here, Roberts and Blazer presented only two years of data, and, although the trial was replicated, did not present any statistical treatment of the data.
FPS 8 produced about 35% lower yield than FPS 1, 3, and 6 (Table 2⇑), in agreement with results of other authors (Bettiga 2003a, Roberts and Blazer 1995). Bettiga (2003a) found that FPS 1, 3, and 6 were not significantly different in yield, which also agrees with the data presented here (Table 2⇑). Roberts and Blazer (1995) stated that FPS 3 produced the highest yields, but without statistical analysis it is difficult to verify that claim. Both Merlot clonal evaluations found that differences in yield were due primarily to variations in cluster weight. Roberts and Blazer (1995) reported that the number of berries per cluster was predominantly responsible for variations in cluster weight, with FPS 8 producing fewer berries per cluster, while Bettiga (2003a) found FPS 8 to produce both fewer berries and smaller berries. Both reports agree with data presented here (Table 2⇑) where FPS 8 did produce both fewer and smaller berries than FPS 1, 3, and 6, although yield differences were predominately due to fewer berries per cluster.
No reports on FPS 9 could be found in the literature. It proved to have the most erratic yield in this trial (Figure 1A⇑). Highly significant interactions of clone by year for yield (Figure 1A⇑) and berries per cluster (Figure 1B⇑) for the total experiment appear largely to be due to erratic performance of FPS 9 alone. In 1997 and 2000, FPS 9 produced as much or more fruit than any other clone, but both of these seasons were followed by low producing years in 1998 and 2001 (Figure 1A⇑). Percentage fruit set was not measured, so it is not clear whether FPS 9 is more sensitive in fruit set to current season’s weather. In any case, the apparent sensitivity of FPS 9 makes it a candidate for research on fruit set but also makes it a questionable choice for growers and vintners looking for consistency in production. Leclair (1995) reported that Merlot is susceptible to poor set in France in some years, but he did not indicate that there was any clonal difference apparent in that characteristic.
Yield and yield components had wide year-to-year variation (Table 2⇑). In many cases, yearly variation exceeded variation among clones. Differences in berry size, although small, did contribute to yield differences, with FPS 8 and 9 having smaller berries than other clones. However, the range in berry size due to clone, a difference of 0.1 g from lowest to highest, was greatly exceeded by the average year-to-year variation in berry size, with a difference of 0.4 g from lowest to highest (Table 2⇑). As with Cabernet Sauvignon (Wolpert et al. 1995), Merlot had only a small difference in berry size due to clone in comparison to yearly variation.
Shoot number differences from year to year were the result of management decisions imposed during shoot thinning before bloom and clearly had an effect on clusters per vine, despite there being seasonal differences in clusters per shoot (Table 2⇑). Consequently, some of the year-to-year differences in yield were due to management decisions.
There was very little clonal variation with respect to fruit composition. It is somewhat surprising that Brix levels are not higher in FPS 8 than in other clones, given its lower yield. Potassium concentration and juice pH were both higher in FPS 8 than other clones (Table 3⇑). Fruit composition did vary significantly from year to year, but as previously noted, that was due to the timing of sampling at harvest.
Vegetative growth as measured by pruning weight was the highest for FPS 8 (1.9 kg per vine) and lowest for FPS 1 (1.5 kg), which agrees with Bettiga (2003a). Yield-to-pruning-weight ratio was lowest for FPS 8, as would be expected for a vigorous and low-yielding clone (Table 2⇑). Correspondingly, FPS 1 had the highest yield-to-pruning-weight ratio (Table 4⇑). Using the ideal ratios of 5 to 10, suggested by Smart and Robinson (1991), clones FPS 1, 3, 6, and 9 were balanced and FPS 8 was excessively vigorous. Vegetative growth was one of the few parameters measured in this trial that did not have significant year-to-year variation. Yearly averages show that this lack of significance does not reflect uniform vigor but, more likely, variation among clones (Table 4⇑).
Conclusion
Clones are often considered for their physical characteristics, such as small berries and loose clusters, as ways to control yield and improve wine quality. However, it is apparent from these results that the environmental differences from year to year exert a greater influence on those yield components than does clonal selection. While growers may wish to avoid an erratic producer such as FPS 9, further investigation into environmental regulation of berry size and fruit set may be as successful a means as choice of clone.
Footnotes
↵5 Current address: Handley Cellars, 3151 Hwy 128, Philo, CA 95466.
Acknowledgments: Authors gratefully acknowledge research funding support from the American Vineyard Foundation.
Thanks are extended to Duckhorn Wine Company for superb cooperation.
- Received August 2005.
- Revision received October 2005.
- Copyright © 2006 by the American Society for Enology and Viticulture
Vol 57 Issue 2








{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
More from this TOC section
Similar Articles
