


Abstract
Trunk diseases diminish vineyard longevity and productivity in nearly every raisin, table, and wine grape production region worldwide. Fungi causing these diseases infect primarily through pruning wounds. One way to control these diseases is to protect pruning wounds with fungicide applications, which can be problematic because of the limited number of registered products; the difficulty for these products to control numerous taxonomically unrelated organisms; the challenge of these products to protect for the entire period of wound susceptibility; and the difficulties and costs associated with hand application of protection treatments. Our goal was to compare the susceptibility of grapevine pruning wounds to various fungi associated with trunk diseases and to evaluate the efficacy of selected fungicides to control these pathogens when applied as pruning wound protectants. The study was conducted over two consecutive years in two separate vineyards in Sonoma and Colusa counties, California. Nine pathogenic fungi were tested: Eutypa lata, Botryosphaeria dothidea, Diplodia seriata, Dothiorella viticola, Lasodiplodia theobromae, Phaeomoniella chlamydospora, Pleurostomophora richardsiae, Togninia minima, and Phaeoacremonium parasiticum. Results showed differences in the infection rates of pruning wounds by these fungi. Species of Botryosphaeriaceae were the most infectious, T. minima, P. parasiticum, P. richardsiae, and E. lata were less infectious, and Pa. chlamydospora was intermediate. Four selected fungicides were tested: 1% Topsin M, Biopaste (5% boric acid in a wound-sealing paste), 1% Cabrio EG, and Garrison. Although results highlight the difficulty of these products to control the entire spectrum of pathogens efficiently, Topsin M was overall the most efficacious product.
Several taxonomically unrelated groups of Ascomycete fungi cause trunk diseases in grapevines: (1) species in the Diatrypaceae family, including Eutypa lata (Moller and Kasimatis 1978), the causal agent of Eutypa dieback; (2) species in the Botryosphaeriaceae, including Botryosphaeria dothidea, Lasiodiplodia theobromae, Diplodia seriata, D. mutila, D. corticola, Dothiorella iberica, D. viticola, Neofusicoccum parvum, N. australe, N. luteum, N. vitifusiforme, and N. viticlavatum (van Niekerk et al. 2004, Úrbez-Torres and Gubler 2009, Úrbez-Torres et al. 2009), fungal taxa that have been reported to cause different grapevine diseases such as Bot canker (Úrbez-Torres et al. 2006, 2008), black dead arm (Lehoczky 1988), and Macrophoma rot (Milholland 1988); (3) species in the Togniniaceae, including Togninia minima (anamorph = Phaeoacremonium aleophilum), Phaeacremonium angustius (Pm. angustius), Pm. mortoniae, Pm. viticola, and Pm. parasiticum (Crous et al. 1996, Dupont et al. 2000, Eskalen et al. 2004, Scheck et al. 1998a); (4) one species in the Herpotrichiellaceae, Phaeomoniella chlamydospora (Pa. chlamydospora) (Crous and Gams 2000); and (5) one species in the Pleurostomataceae, Pleurostromophora richardsiae (Pl. richardsiae) (Halleen et al. 2007). Phaeomoniella chlamydospora, T. minima, and many species of Phaeoacremonium are the causal agents of esca or black measles and Petri disease. Additional taxa are continually being associated with wood cankers and branch dieback worldwide.
Infection of grapevines by these fungal pathogens primarily occurs through pruning wounds. Fungal spores (sexual and asexual) become airborne during and following rain and come in contact with and colonize exposed wood vessels (Eskalen et al. 2007, Petzoldt et al. 1983, Rooney-Latham et al. 2005, Úrbez-Torres and Gubler 2008). Grapevines have the highest risk of infection during the pruning period, from late fall to early spring, because of the high number of wounds made on a single grapevine and the frequency of rain events that occur during that period.
Grapevine wounds remain susceptible to infection by these fungi for several weeks (Eskalen et al. 2007, Munkvold and Marois 1995, Úrbez-Torres and Gubler 2008). Plugging of the xylem and phloem elements and decay of the wood follow infection, impairing translocation of water and nutrients and leading to the decline of the grapevine. Each grapevine can become infected multiple times with one or more pathogens, and it is common to isolate several pathogens from old cankers. Therefore, trunk diseases should be regarded as a complex of pathogens (Larignon and Dubos 1997, Rolshausen et al. 2004, Úrbez-Torres et al. 2006). For these reasons, it is sometimes difficult to associate one type of symptom with a single pathogen. Esca symptoms include the presence of brown to black spots or streaks in xylem vessels, “tiger-striped” chlorotic and necrotic patterns on leaves, and the formation of necrotic spots or “measles” on berries. Wood symptoms of Eutypa dieback and Bot canker are difficult to distinguish, as both diseases exhibit wedge-shape cankers in cross section of infected cordons. Eutypa dieback foliar symptoms typically include the development of stunted shoots and chlorotic and tattered leaves in the spring (Figure 1⇓). No foliar symptoms are known for Bot canker and no characteristic symptoms have been described or identified for the other taxa associated with trunk diseases. Symptom expression also varies from year to year and among grape varieties partly because of variability in susceptibility (Rolshausen et al. 2008, Sosnowski et al. 2007a, 2007b, van Niekerk et al. 2004), making it even more problematic to identify characteristic disease symptoms.

Disease symptoms caused by wood decay fungi in grapevine. A, grapevine cordon dieback with infected spur positions. B, spotting, or measles, on grape berries characteristic of esca disease. C, cross section of grapevine cordon showing wedge-shape canker characteristic of Eutypa dieback or Bot canker. D, cross section of grapevine cordon showing dark vascular streaking characteristic of esca disease. E, stunted shoots and leaves characteristic of Eutypa dieback foliar symptom. F, foliar symptoms characteristic of esca disease.
Grapevine trunk diseases are commonly a significant problem in older vineyards (over 10 years) in every grapegrowing region worldwide. However, young grapevine decline, also called Petri disease or young esca, was also observed in newly planted vineyards (Scheck et al. 1998b). Moreover, Bot canker was observed in California vineyards younger than 10 years (Úrbez-Torres et al. 2008). Once the first symptoms are observed, the affected grapevines usually continue a steady decline until death. In these vineyards, yields are decreased and grape quality is diminished. In addition, the labor involved in pruning out infected wood, retraining dead/dying vines, and replanting adds to overall production costs. When the affected vineyards are no longer economically sustainable to maintain, growers have no alternative but to replant. As a consequence, vineyard longevity can be decreased significantly. It is difficult to accurately measure the economic impact on grape production of trunk diseases because of the slow movement and the long incubation time of some of these fungi. However, losses in California were estimated as $260 million per annum in 1999 (Siebert 2001).
Complete control of these diseases is virtually impossible because of the number of wounds made on an individual grapevine each year and the extended period of wound susceptibility. Moreover, disease pressure is always high in areas where inoculum reservoirs occur on grapevines and on a number of alternate plant hosts. Once the pathogens become established, surgical removal of the diseased wood is critical to reduce inoculum levels. Proper sanitation and cultural practices are essential to delay the establishment and minimize the impact of these diseases. Pruning grapevines in dry weather is critical because fungal airborne inoculum is nearly nonexistent at this time. Late pruning (pruning as close as possible to budbreak) is also strongly recommended because of more rapid wound healing later in the season (Munkvold and Marois 1995). In this regard, double pruning (Weber et al. 2007), which consists of mechanical pruning in the winter followed by hand pruning before budbreak, is a viable technique to complete pruning in large vineyard acreages. Protection of exposed wounds is also highly recommended to achieve good disease control. However, the number of available commercial products that can protect pruning wounds from this broad spectrum of fungi is limited in California. Most products were initially developed and field tested for control of Eutypa lata (John et al. 2005, Moller and Kasimatis 1980, Rolshausen and Gubler 2005, Sosnowski et al. 2008). The two main challenges for registered treatments are to protect pruning wounds against a broad range of taxonomically unrelated pathogens and to persist in wood and provide protection during the entire period of susceptibility without being washed off with rain (Price and Carter 1975, Carter and Price 1977). The goal of our study was to assess the ability of several grapevine pathogens to infect pruning wounds and to test the efficacy of registered and potential products with different active ingredients against these pathogens under field conditions.
Materials and Methods
Field trials were conducted in two separate locations in 2005 and 2006. The Sonoma County site was a 32-year-old Chardonnay vineyard. Grapevines were trained to quadrilateral cordons on a horizontally divided trellis with typically four to six spurs per cordon (16 to 24 spurs per grapevine). The Colusa County site was a 25-year-old Zinfandel vineyard. Grapevines were trained to bilateral cordons with typically five to seven spurs per cordon (10 to 14 spurs per grapevine). Nine fungi known to cause or be associated with grapevine trunk diseases were tested in both locations: Eutypa lata, Botryosphaeria dothidea, Diplodia seriata, Lasiodiplodia theobromae, Dothiorella viticola, Phaeomoniella chlamydospora, Pleurostomophora richardsiae, Phaeoacremonium parasiticum, and Togninia minima. Five pruning wound treatments (i.e., four fungicides plus a water control) were tested against these fungi (Table 1⇓).
Fungicidal pruning wound treatments evaluated in Sonoma and Colusa county field trials, 2005 and 2006.
Grapevine spur positions were pruned to three buds in December at the Colusa County site and in January at the Sonoma County site. For each spur, the paste formulation was painted with a spatula and the liquid formulation was sprayed in a single application with a 100 mL spray bottle. Pruning wounds were artificially inoculated with an aqueous spore suspension of each fungus (~1000 spores per wound) after treatment. Each treatment combination (one pathogen x one fungicide) was applied to every pruning wound made on one grapevine. Treated spurs were allowed to stand for 10 to 11 months and were collected the following November. Spurs were then returned to the laboratory for analyses and isolation.
In 2005, at the Colusa County field trial, 45 treatment combinations (nine pathogens x five fungicides) were applied on 45 grapevines in a completely randomized design. Five replications for each treatment combination were used, for a total of 225 grapevines (45 x 5) for the experiment. Grapevine spurs were artificially inoculated with fungi one day after treatment.
In 2006, at the Colusa County field trial, nine treatment combinations (nine pathogen x one treatment) were applied on nine grapevines. The control treatment was the only treatment collected that year. Five replications for each treatment combination were used, for a total of 45 grapevines (9 x 5) for the experiment. Grapevine spurs were artificially inoculated with fungi seven days after treatment.
In 2006, at the Sonoma County field trial, two sets of data were collected. Forty-five treatment combinations (nine pathogens x five fungicides) were applied on 45 grapevines in a completely randomized design. Three replications for each treatment combination were used, for a total of 135 grapevines (45 x 3) for the experiment. Grapevine spurs were artificially inoculated with fungi one day after treatment on the two cordons trained on one side of the row and one week after treatment on the two cordons trained on the other side of the row. However, for the one-week posttreatment E. lata-inoculated grapevines, only the water-treated spurs (controls) were collected at the end of the season.
The response measured was the percent of one-year-old wood spurs that were colonized with each of the nine fungi evaluated (i.e., the spurs from which the pathogen could be re-isolated). Each treatment combination was calculated as the number of spurs infected with the fungus divided by the total number of spurs inoculated. Fungi were isolated from one-year-old wood in the laboratory as previously described (Rolshausen and Gubler 2005). Wood samples were briefly dipped in ethanol and surface sterilized by flaming before longitudinally split open to expose the vascular system. Five wood chips were sampled from necrotic lesions or vascular discoloration and plated on potato dextrose agar (Difco Laboratories, Detroit, MI) amended with 100 mg/L tetracycline. One culture plate was used per spur sample. After one to three weeks of incubation at room temperature, fungi were identified visually by colony morphology and fungal recovery was calculated for each respective fungus. Spurs were rated as either infected or not infected.
The data are presented as the mean percent infection (MPI) for the two experimental locations over the two years. The mean percent disease control (MPDC) was also presented for each fungicide treatment, where MPDC was calculated on the basis of MPI of the control treatments as (100x (1-(MPI treatment/MPI control))). Data were analyzed using a weighted least square ANOVA with Dunnett’s comparison test to determine if there were significant differences among least square means. All data analysis was performed using SAS software (SAS Institute, Cary, NC).
Results and Discussion
All nine fungi were able to colonize pruning wounds but with differences in infection rates (Figure 2⇓). Interestingly, taxonomically related taxa had similar infection rates. All four Botryosphaeriaceae species tested had the highest infection rate, ranging from 79 to 87%. Phaeoacremonium parasiticum had the lowest infection rate at 20%, and closely related taxa T. minima had an infection rate of 31%. Eutypa lata and Pl. richardsiae had infection rates of 28% and 34%, respectively, similar to the Togniniaceae fungi. Phaeomoniella chlamydospora was intermediate with a 58% infection rate. Pruning wound susceptibility can vary depending on the the age of the pruning wound and the time of pruning. Usually, the susceptibility decreases over time with a faster rate under higher degree days (Munkvold and Marois 1995). Our results supported this tendency, as lower wound colonization was seen in the two trials where grapevine spurs were artificially inoculated with the fungal pathogens one week versus one day after pruning (data not shown). The higher infection rates seen with the four species of Botryosphaeriaceae versus the other fungi tested may reflect a more rapid decrease in pruning wound susceptibility against E. lata, Pa. chlamydospora, Pl. richardsiae, Pm. parasiticum, and T. minima. Additionally, the experiments were carried out with only one isolate per fungal taxon (except for E. lata where ascospores were used). Since intraspecific variation in virulence was previously reported for E. lata (Sosnowski et al. 2007b), the suceptibility of pruning wounds to fungal pathogens may vary depending on the specific isolate used. Under optimal disease conditions, T. minima and Pm. parasiticum produce perithecia and ascospores (Crous and Gams 2000, Rooney-Latham et al. 2005). Further research should examine potential differences in virulence and wound colonization using these other fungal spore types.
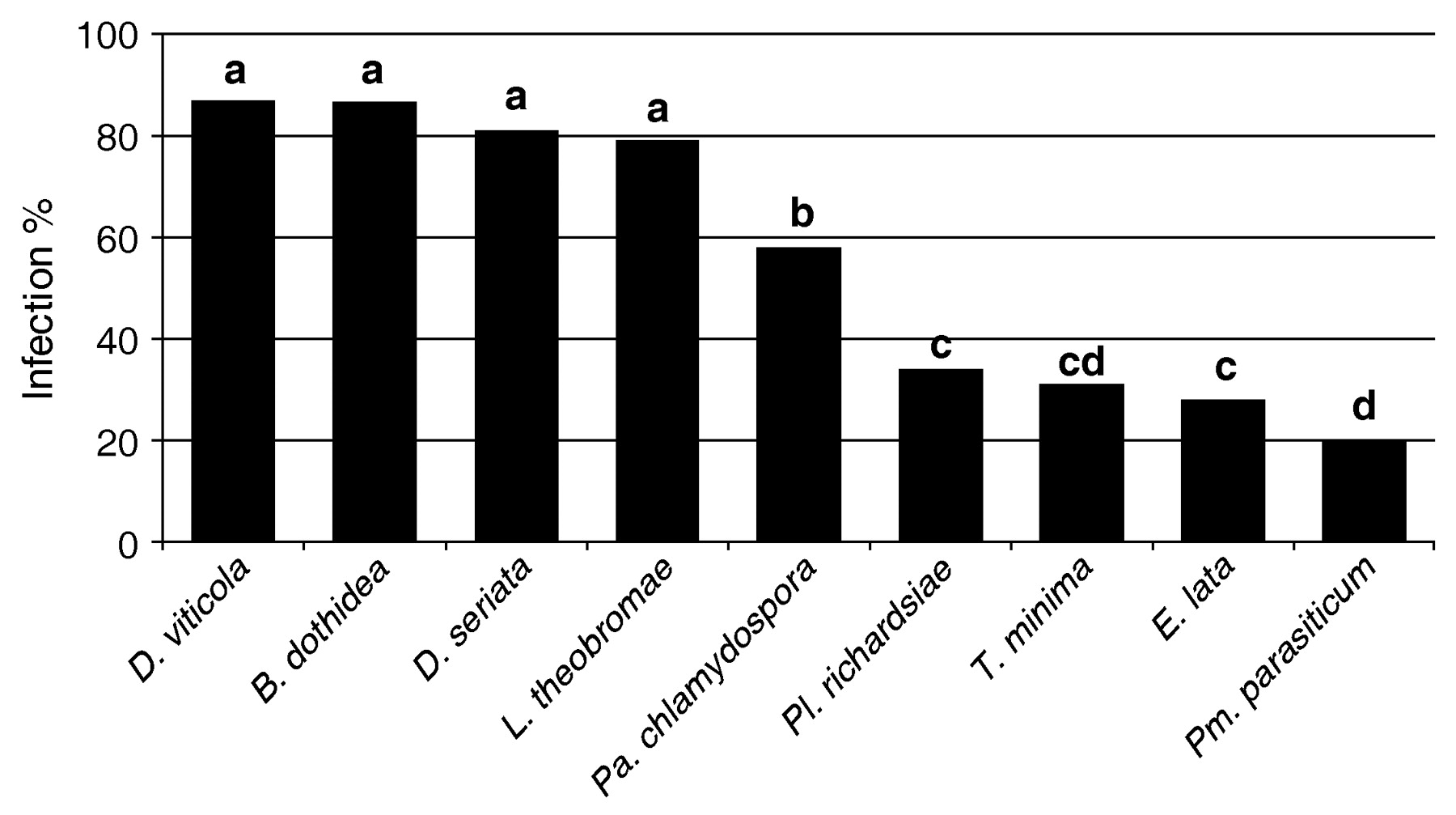
Results of field trials evaluating pruning wound infection rates with nine pathogenic fungi located in two vineyards in Sonoma and Colusa counties, 2005 and 2006. Bars represent the least mean square of percent infection for the two vineyards and for the two years the experiments were conducted. Bars with a different letter are significantly different according to Dunnett’s test (p = 0.05).
Botryosphaeriaceae species have been recently recognized as playing a major role in grapevine wood diseases worldwide and several species have now been identified across California (Úrbez-Torres et al. 2006). Among the four Botryosphaeriaceae species tested here, L. theobromae is more common in the southern San Joaquin Valley (south of Merced County), while B. dothidea is more common in northern California (north of Stanislaus County). Diplodia seriata is ubiquitous throughout California and D. viticola is commonly found in northern and central coastal climates of California. In northern California valley locations (i.e., Napa, Solano, San Joaquin, and Stanislaus counties), the incidence of E. lata in grapevine is typically higher than the Botryosphaeriaceae fungi (Úrbez-Torres et al. 2006). These data highlight the importance of fitness of fungi in disease distribution, incidence, and severity. The establishment of a pathogen in a vineyard depends on several factors, including environmental parameters (temperature, rainfall, surrounding ecosystems), cultural practices (varieties grown, pruning techniques, trellising systems), and pathogen fitness (such as asexual or sexual reproduction and virulence). The ability of fungi to colonize pruning wounds, as measured in this study particularly with regard to Botryosphaeriaceae species, does not automatically translate into high disease incidence in natural vineyard settings. For example, L. theobromae is not a common pathogen in Sonoma and Colusa counties where these experiments were conducted, yet it was able to infect a high proportion of pruning wounds under the natural conditions occurring in these vineyards. Thus, results also highlight the potential threat of these pathogens in production vineyards if they become established in an area.
Protection of pruning wounds is critical to limit infection and subsequent spread of grapevine trunk diseases. Dissemination of T. minima and Pa. chlamyospora through contaminated propagation plant material has also been proposed (Edwards and Pascoe 2004, Fourie and Halleen 2004). Management strategies should be adopted in the nursery to limit the spread in propagation wood, including pruning wound protection of mother vines (Halleen et al. 2007). In California, Benlate (50% a.i. benomyl; DuPont Wilmington, DE) was registered for 30 years as a pruning wound protectant for the control of E. lata. However, the product was removed from the market in 2001, leaving growers with no alternative treatments. To date, there are few treatments currently available for California growers to control trunk diseases, most of which were tested against E. lata. The four products tested in our field trials were originally selected because all contain different active ingredients with different modes of action and all showed some efficacy against E. lata in previous studies (Rolshausen and Gubler 2005, Sosnowski et al. 2008). Benlate was used in a recent study because it was the industry standard for many years (Sosnowski et al. 2008). Our study used Topsin M because it was recently registered on grapevine in California for the control of grapevine wood diseases. Topsin M and Benlate are both systemic fungicides of the methyl benzimidazole carbamates group and have the same mode of action. These compounds impair cell division by targeting the β-tubulin assembly during mitosis. Cabrio EG belongs to the class of strobilurin fungicides that disrupt cell respiration by targeting cytochrome bc. The fungicidal paint Garrison has two active ingredients: iodocarp targets lipids and cell membrane permeability and cyproconazole targets sterol biosynthesis in cell membranes. Boron, which is the active ingredient of Biopaste, is thought to inhibit glycolysis in fungi. Our results indicate that Biopaste, Garrison, and Topsin M provide excellent control of E. lata (Table 2⇓), concurring with results from previous studies (Rolshausen and Gubler 2005, Sosnowski et al. 2008). Although Cabrio EG was the least efficacious product to control E. lata, it still performed statistically better than the water control treatment. Elsewhere Cabrio EG reduced infection by E. lata, but to a lesser extent than Benlate (Sosnowski et al. 2008).
Results of field trials evaluating disease control conducted in two vineyards in Sonoma and Colusa counties, 2005 and 2006. Values represent the least square means for percent infection of pruning wounds treated with four fungicides plus a water control and artificially inoculated with nine fungal pathogens. Results are presented as the average for the locations over both years. Values followed by a different letter in each row were significantly different according to Dunnett’s test (p = 0.05).
Results highlight the difficulty of using chemical treatments to control with the same efficacy a broad spectrum of taxonomically unrelated fungi (Table 2⇑). For example, Biopaste was very efficient in controlling E. lata, Pa. chlamydospora, Pl. richardsiae, and the Togniniaceae fungi but did not perform as well against the Botryosphaeriaceae. Comparable control levels against the Botryosphaeriaceae were obtained with Garrison as those obtained with Biopaste. However, protection levels of Garrison were more erratic against the other fungi, ranging from very good against Pl. richardsiae and E. lata to poor against Pm. parasiticum and T. minima. Cabrio EG was an effective fungicide against the Botryosphaeriaceae group, but was the least effective fungicide against the other taxa. Statistically, the best overall product was Topsin M, perhaps because its mode of action targets a cellular function (mitosis) that is highly conserved across Ascomycetes. In our study, Topsin M was not challenged over time. Fungal inoculations were performed one and seven days after treating the wounds with fungicides. However, pruning wounds can be susceptible to infection for several weeks (Eskalen et al. 2007, Munkvold and Marois 1995, Úrbez-Torres and Gubler 2008). The main limitation of liquid fungicide formulations such as Topsin M is that they usually do not offer full protection for the entire period of susceptibility of pruning wounds (Price and Carter 1975, Carter and Price 1977, Rolshausen and Gubler 2005). Often these liquid formulations are easily washed off with rainfall. Furthermore, the climatic conditions of the two vineyard locations in which the trials were conducted are representative of winter conditions in northern California, but not of other grapegrowing locations. Fungicide and fungi could behave differently on pruning wound surfaces in other climatic environments. Further research should test promising fungicides candidates such as Topsin M against the fungal pathogens at several interval times following application and in different geographical locations to ensure protective efficacy. Moreover, the fungicides tested here were all applied directly on pruning cuts. In reality, paintbrush application of pruning wound protectants is economically feasible only in high-value vineyards. The large acreage combined with the number of pruning cuts on individual grapevines makes it economically and time challenging for growers to protect their vineyard in this manner. Future research should address alternatives. We are currently evaluating a combination of pruning techniques and spray programs on control of these grapevine trunk diseases.
Conclusion
The purpose of this study was to examine the susceptibility of grapevine pruning wounds to nine fungi (Eutypa lata, Botryosphaeria dothidea, Diplodia seriata, Dothiorella viticola, Lasodiplodia theobromae, Phaeomoniella chlamydospora, Pleurostomophora richardsiae, Togninia minima, and Phaeoacremonium parasiticum) associated with trunk diseases and to evaluate the efficacy of four selected fungicides (Topsin M, Biopaste, Cabrio EG, and Garrison) to control these pathogens. Results showed that all fungi were able to infect pruning wounds but with a disparity in infection rates. Results also highlighted the difficulty of chemical treatments to control with the same efficiency a broad spectrum of taxonomically unrelated fungi. Statistically, Topsin M was the best overall product against all these pathogens.
Footnotes
Acknowledgments: The authors acknowledge the American Vineyard Foundation, the USDA Viticulture Consortium West, and the California Competitive Grant Program for Research in Viticulture and Enology for partial funding of this research.
The authors thank Sutter Home Winery and Constellation Wines, U.S., River Oaks Vineyards for vineyard sites.
- Received July 2009.
- Revision received November 2009.
- Accepted November 2009.
- Published online March 2010
- Copyright © 2010 by the American Society for Enology and Viticulture
Literature Cited
Vol 61 Issue 1








{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
More from this TOC section
Similar Articles
