


Abstract
Sangiovese vines mechanically spur-pruned during dormancy in February were manually finished either immediately or post budburst to test the potential of a ‘double-pruning’ approach to delay fruit sugar accumulation and limit yield. The treatments were applied in 2014, 2015, and 2016 at BBCH-0 as standard hand-finishing on dormant buds (SHF), and as late (LHF) and very late (VLHF) hand-finishing at BBCH-14 and BBCH-19, i.e., when the two apical shoots on the mechanically-shortened canes were ~10 and 20 cm long, respectively. While yield per vine was drastically reduced in the VLHF treatment (−43% versus SHF) due to high incidence of unsprouted (blind) nodes, lower shoot fruitfulness, and berries per cluster, yield reduction in LHF was −22% versus SHF due only to the incidence of unsprouted nodes. While the fruit ripening profile was not significantly modified in VLHF compared to SHF, in data pooled over three seasons, LHF delayed basic fruit composition at harvest, producing fruit with less total soluble solids, lower pH, and greater acidity, but more phenolics than SHF. Overall, LHF proved to be effective at reducing yield per vine to a level that did not require expensive cluster thinning. By reducing berry sugar accumulation, it has the potential to produce wines with lower alcohol and higher phenol content. Noteworthy too is its potential to delay harvest date or increase crop hanging time under specific vineyard conditions.
Increased heat accumulation associated with global warming poses a challenge in several grapegrowing areas worldwide (Jones et al. 2005). Attendant weather events include heat waves, drought, and intense rainfall; the agriculture industry and its vineyard sector must devise measures to protect against, offset, or adapt to them, now and in the future (Keller 2010, Palliotti et al. 2014). Crop management practices must therefore be able to rely on more reliable weather models to forecast impending harmful events and on field tools developed through research to cope with them (Palliotti et al. 2013a, 2013b, 2014, Poni et al. 2013, Varela et al. 2015, Gatti et al. 2016a, Paciello et al. 2017).
For instance, the especially hot and dry 2003, 2007, 2009, 2011, 2012, and 2015 seasons in southern Europe showed that one of the most recurring adverse effects in grapevine is the compression of all phenological stages (Mosedale et al. 2016). This trend tends to accelerate ripening stages from veraison in the hotter portion of the growing season, often inducing rapid sugar accumulation, high pH, low acidity, and poor aromatic and phenolic content (Petrie and Sadras 2008, De Orduňa 2010, Palliotti et al. 2014).
In cool climate viticulture districts, late winter pruning is often used to delay budburst as a way of protecting the crop against damage from spring frost (Howell and Wolpert 1978, Trought et al. 1999). Conversely, when applied in other environments, this technique has induced somewhat different results. The most notable is the per-vine yield increase reported in Grenache (Coombe 1964), Ugni blanc (Bouard 1967), Carignon and Grenache (Vergnes 1981), Cabernet Sauvignon, Gewürztraminer, and Palomino (Whittles 1986), and Merlot (Friend and Trought 2007). The most salient effects produced by this practice on yield components were increased fruit set and cluster weight. This was attributed to flowering occurring at a more favorable climatic period, leading to enhanced fertilization of individual flowers and better seed development (Friend and Trought 2007). The same technique also resulted in reduced vine yield in Zante Currant (El-Zeftawi and Weste 1970), Perlette (Jensen and Dokoozlian 1991), Merlot (Keller and Mills 2007), and Cabernet Sauvignon (McGourty 2010). The lower cropping in these trials was due to reduced berry weight, number of clusters per vine, or berries per cluster. On the other hand, no changes in vine yield under the practice were found in Mataro (Barnes 1958), Chardonnay and Merlot (Weber et al. 2007), or Sauvignon blanc (Trought et al. 2011).
Most recently, dramatic yield loss under the same approach was reported in Sangiovese: up to 55% when applied in late spring (early May) and early summer (June) (Frioni et al. 2016), or up to 28% and 91% when applied after budburst, precisely when the apical shoots of unpruned canes were at the stage of about two and seven unfolded leaves, respectively (Gatti et al. 2016b).
This practice likely induces a severe source limitation due to loss of stored carbohydrate reserves used to support the initial flush of vegetative growth, which is then removed by pruning. Depending on the growth stage reached at pruning time, new shoots already bearing some good source leaves are also eliminated. In fact, delayed winter pruning impacts yield linearly in relation to the amount of leaf area removed (Gatti et al. 2016b). Moreover, in unpruned canes, shoot growth of median and basal nodes is increasingly inhibited by the dominance exerted by more distal apical shoots. Delayed winter spur-pruning performed in one step removes all the ‘green tissue’ formed by the whole bud load on the vines. In some cases, due to factors related to trellis geometry, supporting structure, and care taken in performing summer pruning, late-season canes are not exactly vertical. Their position would tend to weaken the control exerted by apical dominance on the subtending nodes (Gatti et al. 2016b) and, at the timing of delayed winter pruning, the leaf area removed can be greater than that of vertical, well-aligned canes.
The trial we report tested the efficacy of double-pruning on vines that were first mechanically pre-pruned in winter and then manually spur-pruned in spring. The working assumption was that hand follow-up on short vertical canes would result in a source limitation sufficient to reduce yield and delay fruit maturity for wine composition.
Materials and Methods
Plant material, environmental conditions, and experimental design
The trial was carried out in 2014, 2015, and 2016 in a vineyard near Perugia (42°58′N; 12°24′E, elev. 405 m asl) in Umbria (central Italy) on mature Sangiovese grapevines grafted on Millardet et de Grasset 420A rootstock, planted at 2.5 × 1.0 m inter- and intrarow spacing and trained to a vertically shoot-positioned, spur-pruned cordon trellis. The cordon was trained 0.9 m aboveground with three pairs of trellis canopy wires on a canopy wall extending 1.2 m above the permanent cordon.
Five adjacent rows were selected to form a completely randomized block design, with each row as a block. Using a rotary disk pre-pruner (Volentieri-Pellenc Visio TLV20), all rows were subjected to winter mechanical prepruning in late winter (mid-February, Figure 1). The pre-pruner was set to shorten the canes to a uniform length of about six to seven nodes each. Within each row, groups of 25 adjacent vines (sub-replicates) were randomly assigned to the following hand-finishing (HF) treatments: (1) standard (SHF), performed right after mechanical prepruning in mid-February with all buds still dormant; (2) late (LHF), performed when the first two apical shoots on the mechanically-pruned canes were at the phenological stage BBCH-14 (Lorenz et al. 1995), ~10 cm long with three to four unfolded leaves, applied on 14, 28, and 11 April in 2014, 2015, and 2016, respectively; and (3) very late (VLHF), performed when the two apical shoots on the mechanically-pruned canes were at BBCH-19, ~20 cm long with eight to nine unfolded leaves, applied on 30 April 2014, 8 May 2015, and 26 April 2016, respectively (Figure 2).

(A) Mechanical prepruner at work in the spur-pruned cordon-trained Sangiovese vineyard, and (B) vines just after the prepruning treatment in winter.

(A) Detail of hand-finishing (HF) performed on very late HF when the apical shoots growing on preshortened canes were at BBCH-19 (~20 cm long with eight to nine unfolded leaves), and (B) pre- and post-HF comparison.
HF in all treatments consisted of cutting back all canes of the test vines to standard two-node spurs. In all years, local standard practices were applied for pest management, no leaf removal was performed, and shoots were mechanically trimmed, usually once per season, when they were 20 cm above the last set of catch wires to prevent canopy shade being cast over the fruiting zone. Weather conditions were monitored by an automatic A753 GPRSRTU SEN-R meteorological station (Davis Instruments Corp.), located ~600 m from the vineyard.
Leaf area development and vine size
In all years, 16 fruiting shoots per treatment were randomly collected from 16 vines within the experimental blocks just after harvest. Total leaf area per shoot was measured by an AAM-7 leaf area meter (Hayashi-Denko), and total vine leaf area was calculated by multiplying shoot mean leaf area by shoot number per vine. Canes from 16 representative vines per treatment (four vines per block) were weighed yearly after winter pruning to estimate annual vine growth; the resulting data were used to calculate the Ravaz index (yield-to-pruning weight ratio, kg/kg; Ravaz 1903). In LHF and VLHF vines, the pruning weight resulting from winter mechanical pruning was added to the cane weight recorded at HF. Vine balance was measured by calculating the ratio of total leaf area to vine yield in all treatments.
Ripening kinetics, vine yield, and must composition at harvest
In 2015 and 2016, total soluble solids (TSS), titratable acidity (TA), and must pH were periodically analyzed using random samples of three replicates of 100 berries per treatment taken from late July until harvest. Brix and pH were measured with a temperature-compensating RX5000 refractometer (Atago-Co Ltd) and a digital pHM82 pH meter (Radiometer), respectively. A Titrex Universal Potentiometric Titrator (Steroglass S.r.l.) was used to measure TA by titrating with 0.1 N NaOH to an end point of pH 8.2; the results are expressed as g/L tartaric acid equivalents (Famiani et al. 2015).
The LHF treatment exhibited two types of shoot: regular shoots, whose clusters had normal growth and ripening patterns, and delayed shoots, with clusters having less-advanced fruit ripening (Figure 3). The ripening kinetics of clusters developed on these two shoot types was therefore tracked separately by selecting 50 shoots per type. These shoots were taken from uniform vines (10 vines per block) and chosen when the shoots showing normal growth were at the onset of veraison.

Photos taken in 2015 on (A) 3 Aug during veraison, showing clusters on “regular” shoots; and (B) on 21 Aug during fruit ripening, showing clusters on “delayed” shoots.
Harvest dates were 24, 10, and 27 Sept in 2014, 2015, and 2016, respectively. In all years, vines from all treatments were harvested on the same day, i.e., when sugar accumulation, indexed as TSS, in the berries of the SHF treatment reached a plateau, and before any sign of berry shrivel. Grapes from all trial vines were manually picked and crop weight and cluster number per vine recorded. Mean cluster weight, berry fresh weight, and number of berries per cluster were also measured. Total anthocyanin and phenolic concentrations were determined on five replicate samples of 50 berries each per treatment, and their concentration was given as mg per kilo of fresh berry weight as described (Iland et al. 1993).
Carbohydrates and nitrogen storage in permanent vine organs
At the end of January in 2015 and 2016, the soluble sugars, starch, and total nitrogen (N) concentrations in roots (fine brown 1.5 ± 0.2 mm diam, taken at 20 to 30 cm soil depths) and canes (internode between buds three and four) were determined on 10 replicates per treatment by colorimetric method (Loewus 1952).
Each replicate was prepared by mixing three different pieces of internodes from one-year-old canes taken between node three and four from the base. The same procedure was used for root analyses, using three pieces of fine brown root per replicate.
Absorbance readings at 620 nm were performed on a Jasco V-630 spectrophotometer. Total N concentration was determined by titration on the same material after Kjeldahl digestion and expressed on a dry weight basis (%).
Node and shoot fertility
Node and shoot fertility were assessed on 20 uniform vines per treatment (four vines per block) in mid-June every year by counting the total number of inflorescences formed per shoot (shoot fertility) and per node retained on the vines after HF (node fertility).
Microvinification and wine analysis
In 2015, wines were made from the crop of SHF and LHF treatments using a microvinification technique. Wines from the VLHF treatment could not be made because low yields made it unfeasible to get at least two replicates of suitable amounts. At harvest, grapes from 130 SHF and 130 LHF vines were harvested manually and transported to the experimental winery in 20-kg plastic boxes. For each treatment, the total harvested grape mass was divided into two lots, each weighing ~180 kg for the SHF treatment and 140 kg for the LHF treatment. Each lot was mechanically crushed, destemmed, transferred to 100-L stainless-steel fermentation containers, sulfited with 35 mg/L SO2, and inoculated with 35 mg/L of a commercial yeast strain (Lalvin EC-1118, Lallemand Inc.). Wines were fermented for 17 days on the skin and punched down twice daily, with the fermentation temperature ranging from 20 to 27°C. After alcoholic fermentation, the wines were pressed at 0 Brix and inoculated with 30 mg/L of Oenococcus oeni (Lalvin Elios 1 MBR, Lallemand Inc.). After completion of malolactic fermentation, the samples were racked and transferred to 60-L steel containers and 25 mg/L SO2 was added. Two months later, the wines were racked again, bottled into 750-mL bottles, and closed with cork stoppers. After one year, the wines were analyzed for alcohol, TA, and pH (Iland et al. 1993). Wine color intensity (OD420nm+OD520nm), color hue (OD420nm/OD520nm), and total phenol and anthocyanin concentrations were determined by spectrophotometer. Total phenols were quantified by measuring the absorbance at 280 nm wine diluted 1:100 with distilled water as described (Ribéreau-Gayon 1970). Anthocyanins were analyzed as reported (Ribéreau-Gayon and Stonestreet 1965). All determinations were carried out in duplicate.
Statistical analysis
A two-way analysis of variance (ANOVA) on the complete randomized block design was used to analyze the timing of HF and year effects on yield components, grape composition at harvest, leaf area development, and reserve storage in canes and roots. ANOVA was performed with blocks as replications (degree of freedom = 4). Mean separation was performed using the Student-Newman-Keuls test (p ≤ 0.05 and 0.01). When a significant Y × T was found, it was partitioned over single year × treatment combinations. Wine parameters were analyzed using one-way ANOVA and differences were determined using the t-test. All the statistical analyses were performed with the SigmaStat 3.5 software package (Systat Software Inc.). Results of the seasonal evolution of TSS and TA are shown as means ± standard error (SE).
Results
Environmental conditions
Heat accumulation, expressed as growing degree days (GDD, calculated with a base temperature of 10°C from 1 April to 30 Sept), was greater in 2015 than in 2014 or 2016 (2034 GDD instead of 1558 and 1848, respectively). Total 2015 rainfall was only 218 mm, rather than 508 or 458 mm in 2014 or 2016 (Figure 4). In 2014, rainfall was uniformly distributed from April to September, but was concentrated in August in 2015, with 105 mm, and heavy from late April to June in 2016. Summer 2015 was marked by high daily maximum air temperatures, reaching 32°C in May, 34°C in June, and 40°C in July and August (Figure 4). In 2014 and 2016, the max air temperature never exceeded 35 and 37°C, respectively. Despite such trends and the absence of irrigation, no visual symptoms of water stress or significant leaf yellowing were observed in the 2015 season.
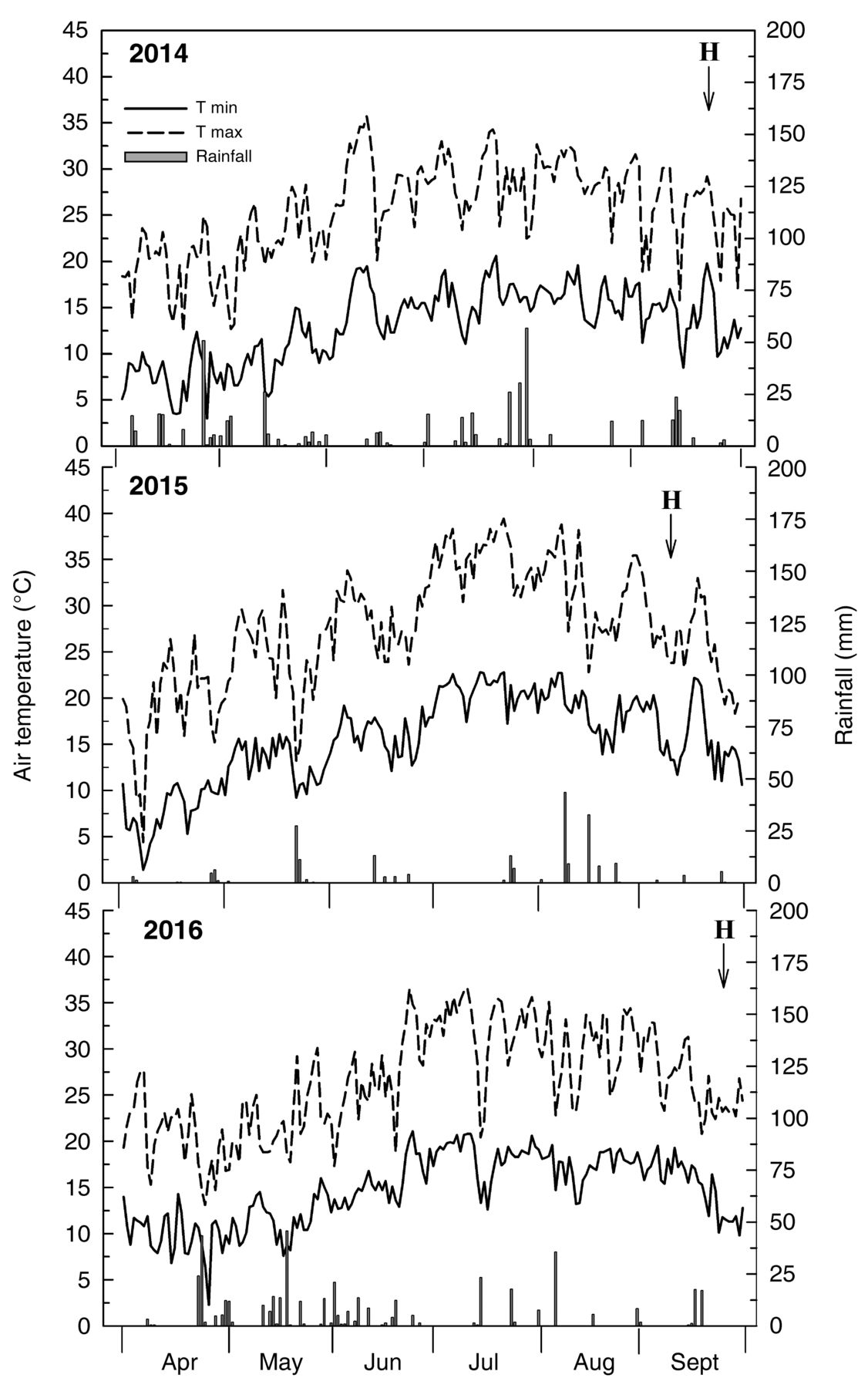
Seasonal trends of maximum and minimum air temperature (T) and rainfall in 2014, 2015, and 2016 seasons. H indicates harvest date.
Vine phenology
Irrespective of treatment and year, all vines were mechanically pre-pruned in mid-February to retain 15.4 ± 1.7 canes on average and ~100 to 110 nodes per vine (each pruned cane had about seven nodes). In all years, SHF vines reached budburst during the third week of March, while budburst in LHF and VLHF was postponed by about two and four weeks, respectively (Table 1). When distal shoots developing along the canes of LHF reached the three to four unfolded leaves stage (BBCH-14), the two basal count nodes ranged from dormancy (BBCH-0) to budburst (BBCH-08). Conversely, at the time of HF, the two basal nodes (retained as spurs) in VLHF vines were in a more advanced stage of growth, ranging from budburst (BBCH-8) to four unfolded leaves (BBCH-14). However, for data pooled over seasons, ~20% of basal buds (first two buds) in LHF and VLHF vines stayed dormant, compared to only 5% assessed in SHF vines (Table 2).
Phenological stage dates for spur-pruned Sangiovese vines mechanically prepruned in February and subjected to hand-finishing (HF) at different times: in February (SHF, standard HF) and after budburst, when apical shoots on canes retained during prepruning were ~10 cm long (LHF, late HF) or ~20 cm long (VLHF, very late HF). Italics indicate the dates of flowering and veraison on “delayed shoots.”
Crop weight and yield components at harvest in 2014, 2015, and 2016 on spur-pruned Sangiovese vines mechanically prepruned in February and subjected to hand-finishing (HF) at different times: February (SHF, standard HF) and after budburst, when apical shoots on canes retained with prepruning were ~10 cm long (LHF, late HF) or ~20 cm long (VLHF, very late HF).
The phenology difference among treatments at budburst was broad and ranged between +24 days in 2016 and +29 days in 2014 (Table 1). The delay narrowed at bloom to +19 days and was further reduced to +7 to 10 days at veraison. This pattern pertained to the ‘regular shoots’ derived from the buds that at the time of HF were at least at BBCH-7 (beginning of budburst with green shoot tips just visible). In LHF vines, all buds between BBCH-1 and BBCH-7 (~20 to 25% of the total buds on the vines after HF) produced ‘delayed shoots’ with flowering postponed by ~15 days in comparison to the ‘regular shoots’ arising from the same vines, i.e., 12 June versus 28 May in 2015 and 10 June versus 1 June in 2016 (Table 1). Unlike LHF vines, the VLHF never exhibited these two shoot types, probably because over the approximate two weeks from BBCH-14 to 19, the two basal buds of the spurs burst (from BBCH-8 forward), except for the 22% of nodes that remained dormant (Table 2).
Effects of delayed HF on yield components, ripening kinetics, and grape composition
LHF and VLHF had 20% fewer shoots per vine than SHF (Table 2). VLHF also had significantly fewer clusters per shoot. No significant T × Y interaction was found for any of the yield component parameters reported in Table 2.
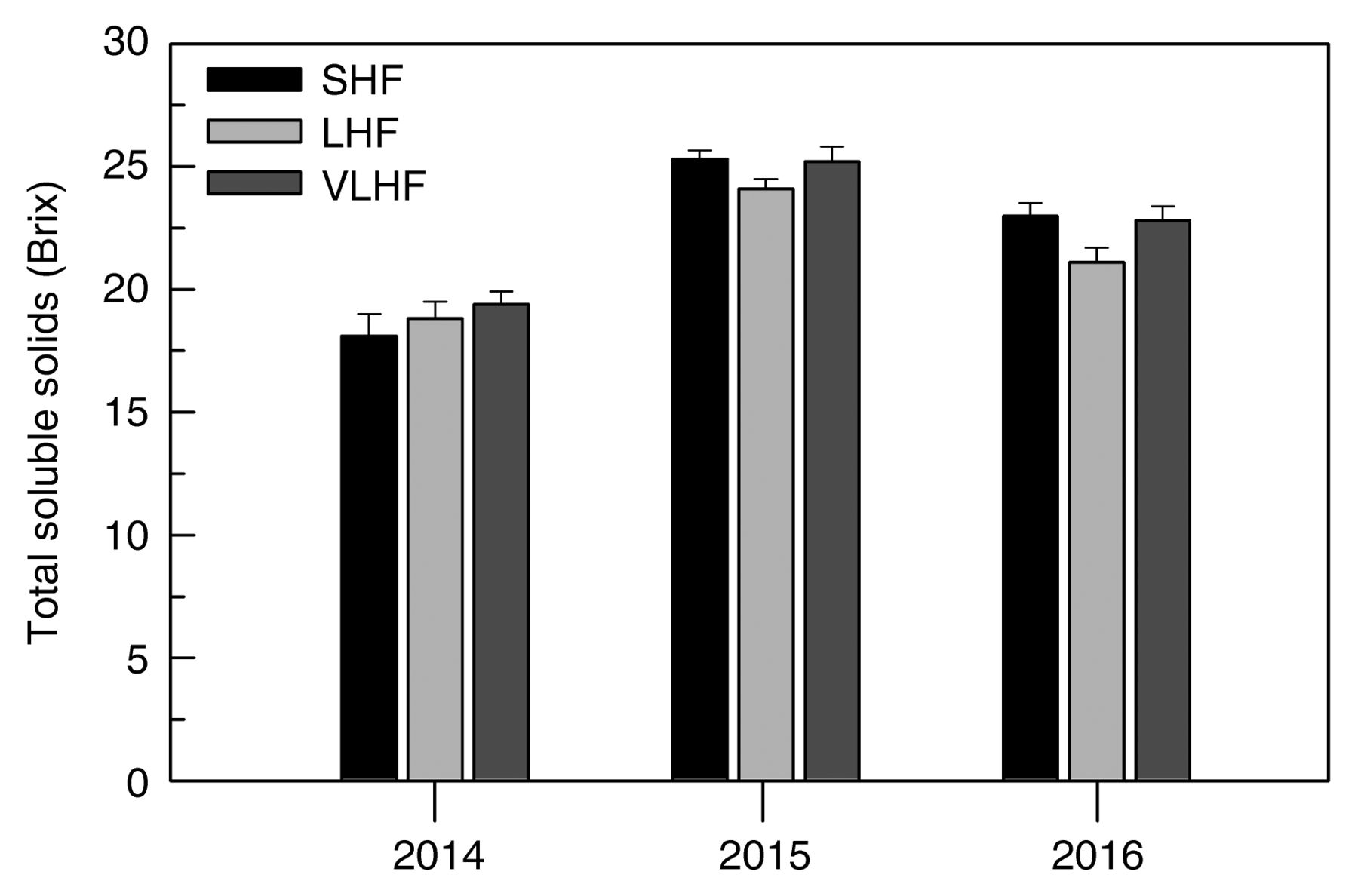
Pooling the data across trial seasons showed that LHF and VLHF vines had lower yields per vine (22% and 43%) than SHF (Table 2). In LHF, this was primarily due to a 20% reduction in clusters per vine, while all yield components were affected in VLHF: ~39% reduction in clusters per vine, reduced cluster weight (15%), and 16% fewer berries per cluster. Total must soluble solids concentration at harvest was significantly lower in LHF than in SHF and VLHF, but TA increased and must pH was lower (Table 3). An increase in phenol concentration, ranging from 18 to 20%, was recorded in LHF and VLHF; total anthocyanins were unaffected. However, TSS showed a significant T × Y interaction that, when partitioned (Figure 5), showed that while in 2014 TSS was generally low in all treatments, in 2015 and, to a greater extent, in 2016, TSS was notably lower in LHF than in SHF and VLHF.
Fruit composition at harvest in 2014, 2015, and 2016 on spur-pruned Sangiovese vines mechanically prepruned in February and subjected to hand-finishing (HF) at different times: February (SHF, standard HF) and after budburst, when apical shoots on canes retained with prepruning were ~10 cm long (LHF, late HF) or ~20 cm long (VLHF, very late HF).

Interactive treatment × year effects on total soluble solids at harvest. For each treatment × year combination, vertical bar represents SE (n = 5). SHF, standard hand-finishing, LHF, late hand-finishing, VLHF, very late hand-finishing.
Ripening kinetics showed that sugar accumulation and juice TA degradation were reduced in clusters on delayed shoots compared to regular shoots on the same vines and to SHF and VLHF vines (Figure 6).
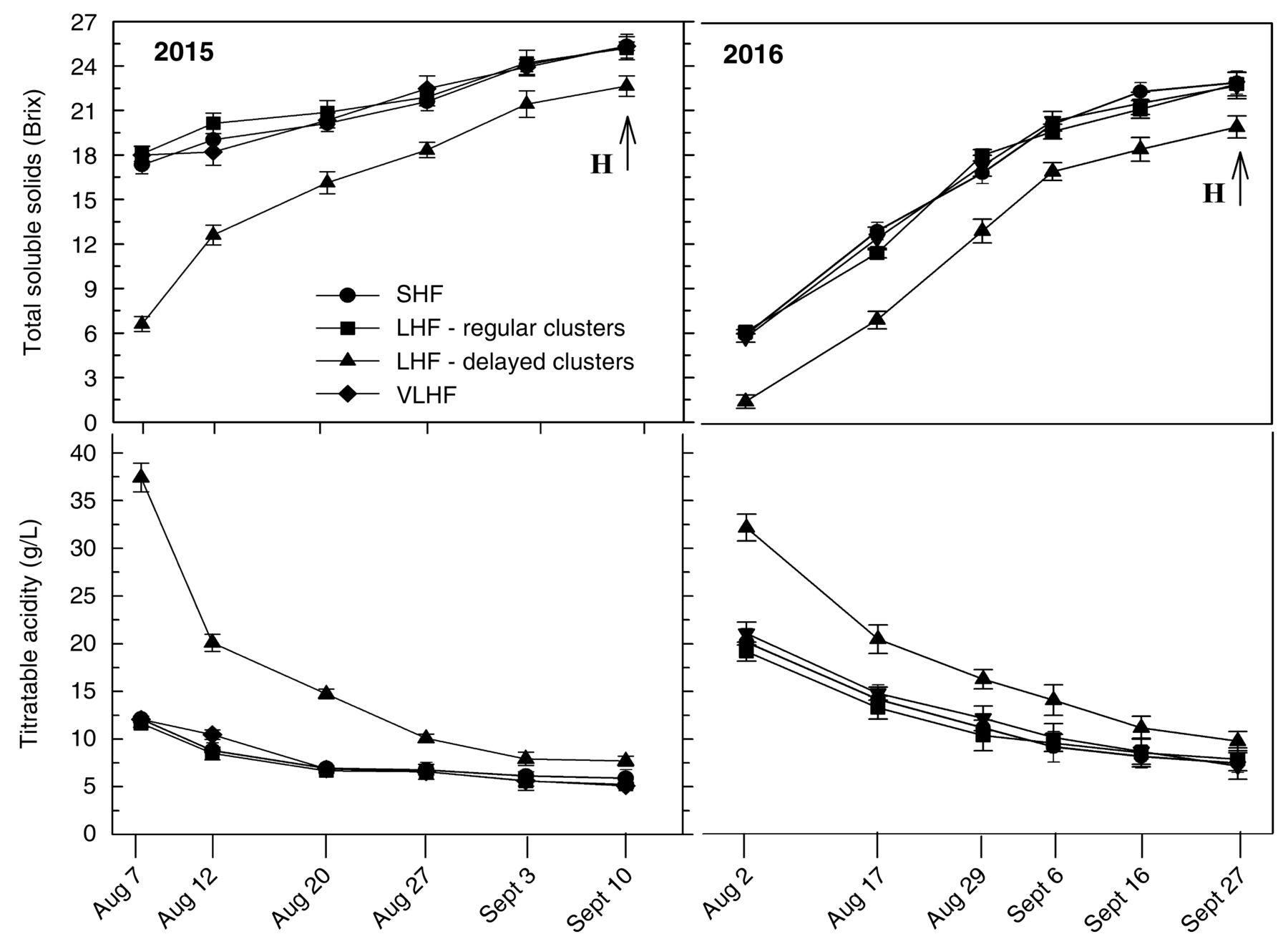
Seasonal changes of total soluble solids and titratable acidity in 2015 and 2016 on spur-pruned Sangiovese vines mechanically prepruned in February and subjected to hand-finishing (HF) at different times: February (SHF, standard HF) and after budburst, when apical shoots on canes retained with prepruning operation were ~10 cm long (LHF, late HF) or ~20 cm long (VLHF, very late HF). Data points are means of three replicates of 50-berry samples. Vertical bars represent SE. H indicates harvest date.
Comparing seasons indicates that the warmer 2015 led to a significant decrease of yield due to fewer clusters per vine and lighter weight of clusters and berries (Table 2); higher TSS, anthocyanins and phenols, and lower TA were found than in 2014 and 2016 (Table 3).
Vegetative growth, vine size, and carbohydrate reserves
Estimated total leaf area per vine (data pooled over seasons) removed with the HF was 0.21 m2 for LHF and 0.43 m2 for VLHF (Table 4). There were no differences in total leaf area and lateral leaf area per vine or one-year-old pruning weight across treatments at the end of vegetative growth. Likewise, no significant T × Y interactions were found for these parameters. Due to their lower yield, the VLHF vines had a significantly greater leaf-to-fruit ratio and lower yield-to-pruning weight ratio than the other treatments (Table 4). In 2014, the leaf-to-fruit ratio was reduced in comparison to the other years, while the yield/pruning weight ratio was increased by ~25% (Table 4). In 2015 and 2016, soluble sugars, starch, and total N contents in canes and roots at the end of January did not vary across treatments (Table 5).
Leaf area characteristics, winter pruning weight, and balance indices in 2014, 2015, and 2016 for spur-pruned Sangiovese vines mechanically prepruned in February and subjected to hand-finishing (HF) at different times: February (SHF, standard HF) and after bud-burst, when apical shoots on canes retained with prepruning were ~10 cm long (LHF, late HF) or ~20 cm long (VLHF, very late HF).
Cane and root reserves and total nitrogen at the end of January 2015 and 2016 on spur-pruned Sangiovese vines mechanically prepruned in February 2014 and subjected to hand-finishing (HF) at different times: February (SHF, standard HF) and after budburst, when apical shoots on canes retained with prepruning were ~10 cm long (LHF, late HF) or ~20 cm long (VLHF, very late HF). DW, dry weight.
Wine characteristics
Wines made from LHF grapes after one year of aging had 0.7% less alcohol than wines made from SHF grapes; total acidity, pH, total dry extract, and anthocyanins were similar (Table 6). The concentration of phenols was instead significantly increased in the LHF wines (+18%), while chromatic intensity and color hue of the wines were unaffected.
Wine composition in 2015 in spur-pruned Sangiovese vines mechanically prepruned in February 2014 and 2015 and subjected to hand-finishing (HF) at different times: February (SHF, standard HF) and after budburst, when apical shoots on canes retained with prepruning were ~10 cm long (LHF, late HF). Grape batches from very late HF could not be vinified due to too-low yield per vine.
Node fertility
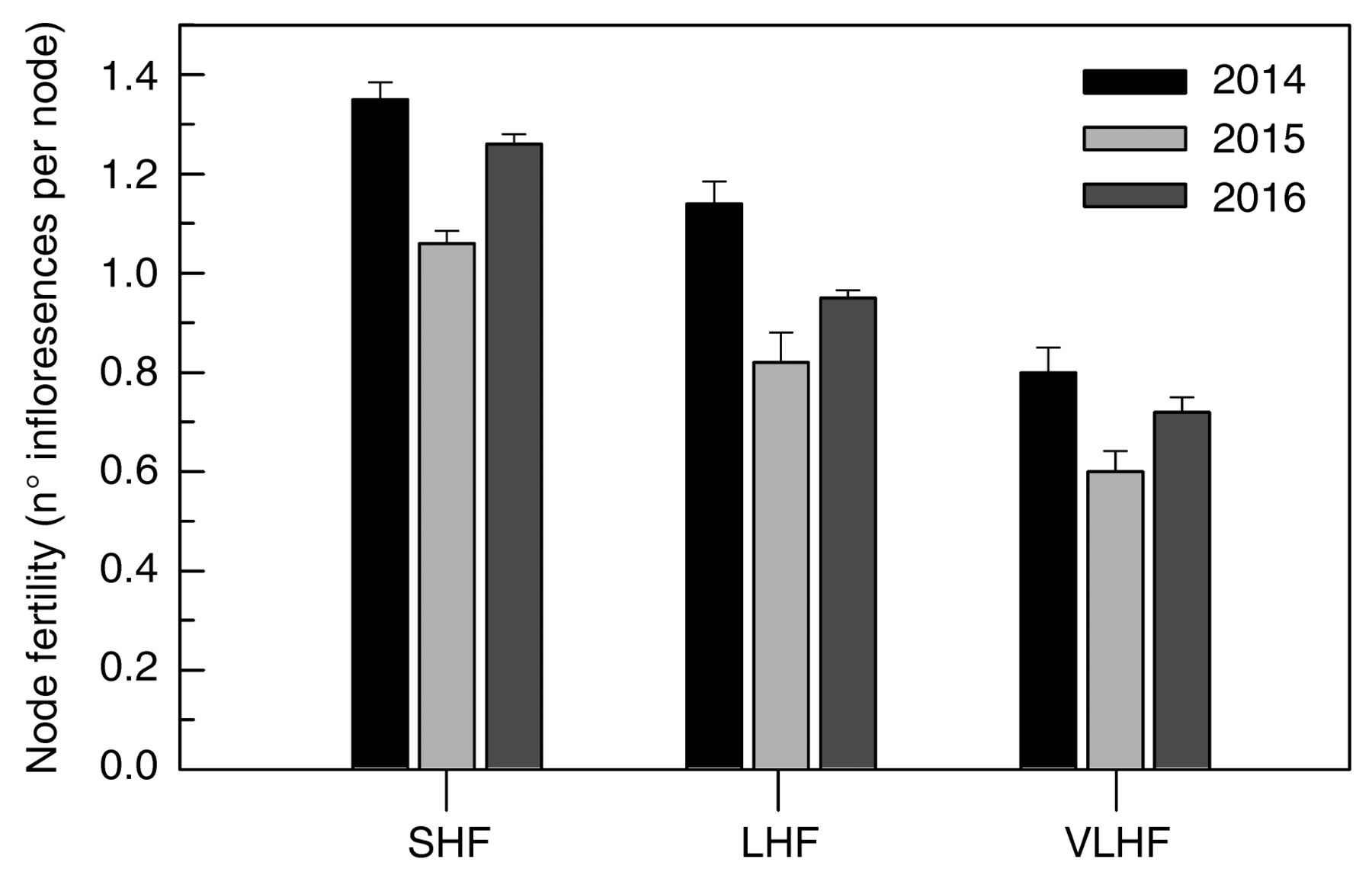
Node fertility, including contributions of both primary and secondary shoots, was significantly lower across years in LHF than in SHF (i.e., −16% in 2014, −23% in 2015, and −25% in 2016) (Figure 7). A greater reduction in node fertility was recorded in the VLHF treatment (−41% in 2014 and −43% in 2015 and 2016). Regardless of treatment, a lower 2015 node fertility was assessed, scoring ~0.2 to 0.3 fewer inflorescences per node compared to the other years (Figure 7).

Node fertility in 2014, 2015, and 2016 of spur-pruned Sangiovese vines mechanically prepruned in February and subjected to handfinishing (HF) at different times: February (SHF, standard HF) and after budburst, when apical shoots on canes retained with prepruning operation were ~10 cm long (LHF, late HF) or ~20 cm long (VLHF, very late HF). Each bar is the mean of 20 vines ± SE.
Discussion
Postponing spur-pruning of cordon-trained vines until well after winter mechanical pruning delayed budburst, bloom, and veraison.
When compared to vines subjected to standard winter pruning (i.e., mechanical prepruning immediately followed by HF), the phenology difference was greatest at budburst (up to four weeks), thereafter decreasing to less than three weeks and one week from flowering and veraison, respectively. The extent of delay is related to two factors: timing of HF and the length of the canes retained by mechanical prepruning. It is known that the later HF is performed, the more active leaf area is removed. A negative linear relationship was found between leaf area per vine removed by delayed winter pruning and yield per vine (Gatti et al. 2016b). No more than 10% of the final leaf area should be removed to prevent excessive yield reduction and, hence, make the technique economically unsustainable. Concurrently, if yield is strongly limited, ripening is also accelerated and contributes to offsetting the initial phenological delay. The same authors also show that a significant delay in budburst will prompt shoot growth to shift to a warmer period; consequently, less time is needed for leaves to reach full expansion, due to better environmental conditions with the potential for higher leaf photosynthetic rates. These factors may explain the progressive seasonal reduction in the initial phenology delay.
LHF at BBCH-14 induced a significant reduction in yield per vine by ~22% due to fewer shoots/vine, leading in turn to fewer clusters per vine (Table 2). Interestingly, cluster number per shoot and berries per cluster were not different from standard pruning, suggesting that the increase of the fraction of spur buds that stayed dormant accounted for most of the yield reduction (Table 2). Flower formation is negatively affected by alterations in the photosynthetic pool during the same year, while perturbation of reserve replenishment during the previous year affects bud induction and initiation of the inflorescences (Lebon et al. 2008). LHF had no significant variation in carbohydrates and total N concentration in roots and canes, suggesting that potential fruitfulness of the dormant bud during year one of the bud cycle was not impaired (May 2004, Bennett et al. 2005). Concurrently, that neither clusters per shoot nor berry number per cluster was affected suggests that the number and ‘structure’ of the formed inflorescences in LHF were unaffected by the source limitation induced by HF.
It remains to be seen why LHF showed a higher proportion of non-sprouted nodes (Table 2), detracting from potential yield. The same response was not reported on the same cultivar with a very similar stage of delayed pruning (about two unfolded leaves on apical shoots), although this study used unpruned canes having at least 17 to 20 nodes (Gatti et al. 2016b). In theory, the two basal nodes in our study that eventually formed the fruiting spur on mechanically pre-shortened canes should be less inhibited than the same nodes borne on unpruned canes. However, most of the nodes overlying the two basal ones in our pre-shortened canes had developed into active growth (Figure 2). The theory of acrotony on a vertical cane implies that inhibition of median and basal nodes is induced by a progressive build-up of downward-translocated auxins, primarily synthesized by young tips and unfolding leaves, that becomes a growth-limiting factor beyond a given threshold (Bangerth 1989). It is therefore likely that in pre-shortened canes, either due to the shorter distance auxins have to travel from top to bottom of the cane, or to the higher fraction of growing shoots over total than in unpruned canes, the inhibition of basal bud growth exerted by auxins is more severe, resulting in more nodes that remain dormant.
For the same reason, the VLHF treatment also had a high fraction of non-sprouted basal nodes, though its yield loss was mostly due to lower shoot fruitfulness (0.79 versus 1.09 clusters/shoot) and fewer berries/cluster (93 versus 110) than SHF’s. A non-limiting source-to-sink ratio by the time of compound bud induction is historically acknowledged as a primary regulator of fruitfulness (Coombe 1962). Since VLHF’s leaf area-to-yield ratio was greater than SHF (Table 4), and it showed no differences in reserves storage in winter (Table 5), there is no reason to think that inflorescence induction was inherently limited in this treatment. This indicates that attention should focus on those factors inhibiting the conversion of inflorescence primordia to complete inflorescences.
This process resumes as shoot development begins in the spring during the second season of the bud differentiation cycle (Carmo Vasconcelos et al. 2009) and continues for 15 to 20 days after budburst in Grenache and Merlot (Bernard and Chaliés 1987). Thus, the severe source limitation caused by VLHF (0.43 m2 of LA removed with HF) may have caused pre-developed inflorescence primordia to revert into tendrils. Previous studies reported that this phenomenon is indeed possible and phosphorous supply (Skinner and Matthews 1989), early water deficit (Matthews and Anderson 1989), and application of exogenous gibberellin (Yahyaoui et al. 1998) are among the causal factors involved.
The second yield-limiting mechanism recorded in VLHF, fewer berries per cluster at harvest, may stem from a chain of factors involving both determination of initial flower number per cluster and fruit set, which in turn depend on others (May 2004). For example, a lack of sugar availability in flowers at female meiosis may lead to flower abortion (Lebon et al. 2008). A much-delayed budburst causes shoot growth to occur at higher daily rates under higher air temperatures, thereby exerting stronger competition among the differentiating bud meristems (May 2004). Non-structural cane and root carbohydrates in winter were unaffected by the timing of HF (Trought et al. 2011, Frioni et al. 2016). This suggests that carbohydrate demand post-fruit set did not exceed source availability, allowing normal replenishment of stored reserves.
The formation in LHF vines of two types of shoots, characterized by clusters that follow different ripening kinetics on the same vine, was responsible for lower sugar accumulation and must pH, higher TA at harvest, and lower wine alcohol content. In our LHF vines, ~25 to 30% had a drastic delay in the onset of fruit ripening behind shoots with the regular ripening pattern. Spur buds at stages between bud swell (BBCH-01) and onset of budburst (BBCH-07) at the time of HF (on average 2.5 ± 0.6 buds per vine) originated delayed shoots, while those transiting from budburst (BBCH-07) and beyond originated normal shoots, quantified as 9.5 ± 1.2 buds per vine. This finds indirect support in the lack of delayed shoots on VLHF vines, where HF and consequent leaf removal, which occurred two weeks later than in LHF, coincided with bud development beyond the BBCH-07 phase. Thus, the initial stages of bud development (BBCH-01 to 07) seem to be very sensitive to any coincident source limitation. However, we found that this sensitivity did not result in more conversion into tendrils or limit flower differentiation (Table 2), rather, it resulted in a notable delay of vegetative and reproductive development (Table 2). The causes of this response need to be elucidated. The increase in total phenols in LHF vines could be a consequence of yield containment, since no significant variations occurred in leaf-to-fruit and yield/pruning weight ratios.
In general, very few T × Y interactions were found for all parameters, indicating that effects due to the type of pruning regime were quite stable across seasons. A partial exception was TSS, which in the cool and wet 2014 season was slightly higher in the “delayed” HF treatments versus SHF, while over the two following seasons, LHF manifested and strengthened its delaying effect. While a season climatically not conducive to high photosynthesis, hence high sugar accumulation, might have dampened differences among treatments, the increasing ability of LHF to function as a delaying technique could indicate that some time is needed for the vine to adjust to the new technique.
Conclusions
While the most delayed VLHF treatment reduced yield an excessive 43% below standard pruning, LHF showed several advantages to the double-pruning approach. Post-budburst deferral of HF does not involve additional costs as compared to standard mechanical winter pruning with follow-on manual finishing. In effect, it is a more psychologically acceptable option than having fully unpruned vines until after vegetation commences. It also delayed ripening for grapes whose wine calls for lower sugar and pH and higher TA, while increasing total phenols for production of wines of lower alcohol content, inducing moderate reduction of per-vine yield and extending the harvest window. The latter is a benefit that may be very useful under high acreage of the same or similar cultivars marked by contemporary ripening. If supported in trials of other varieties at other sites, yield reduction could profitably replace the expensive cluster thinning often needed in high-cropping varieties characterized by high bud fertility and large clusters like Sangiovese, Montepulciano, Ciliegiolo, Trebbiano, or Malvasia. Double-pruning can thus help to mitigate the apparent negative impact of global warming trends on grape composition and wine styles (De Orduňa 2010, Keller 2010, Palliotti et al. 2014, Varela et al. 2015).
- Received February 2017.
- Revision received April 2017.
- Accepted April 2017.
- Published online September 2017
- ©2017 by the American Society for Enology and Viticulture
Literature Cited
Vol 68 Issue 4








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
More from this TOC section
Similar Articles
