


Abstract
Thirty-five strains of lactic acid bacteria (20 species) were screened for the ability to produce volatile phenols from the corresponding phenolic acids, p-coumaric and ferulic, in culture medium. The concentration of vinylphenols and ethylphenols in the growth medium was analyzed by gas chromatography. Results showed that 13 strains (37%) were able to produce volatile phenols from p-coumaric acid, although only three (9%) produced 4-ethylphenol, the final product of the p-coumaric acid metabolic pathway. The reduction step of this pathway was only found in the Lactobacillus genus: L. brevis, L. collinoides, and L. plantarum. Seven of the eight pediococci strains studied were able to produce 4-vinylphenol but not 4-ethylphenol from p-coumaric acid. The two Oenococcus oeni strains and the strain of Leuconostoc mesenteroides studied did not produce either of the two p-coumaric acid derivatives. Strains that produced volatile phenols were used in subsequent studies at lower phenolic acid concentrations. Experiments with added 5 mg/L of p-coumaric acid showed that some strains can still produce relatively high (up to 800 μg/L) concentrations of 4-ethylphenol. The capacity of lactic acid bacteria to produce volatile phenols from ferulic acid was much lower than its capacity to produce volatile phenols from p-coumaric acid.
Volatile phenols are aromatic compounds that, above certain levels, negatively affect wine quality, imparting animal, leather, and “horse sweat” odors. Winemakers consider the presence of these compounds in wine a key concern in the control of wine quality. The precursors of the volatile phenols are natural constituents of grape juice and wine: the hydroxycinnamic acids p-coumaric and ferulic (Heresztyn 1986, Chatonnet et al. 1995). The transformation of these compounds involves a sequence of two enzymatic reactions. In the first, the hydroxycinnamic acid is decarboxylated into the corresponding vinyl derivative (4-vinylphenol from p-coumaric acid or 4-vinylguaiacol from ferulic acid); in the second reaction a reductase converts the vinyl into the corresponding ethyl compound (4-ethylphenol or 4-ethylguaiacol) (Heresztyn 1986).
Which wine microorganisms are involved in the metabolism of the hydroxycinnamic acids has been a subject of intense discussion. The ability of Brettanomyces, specifically the species B. bruxellensis and B. anomalus, and its ascosporogenous form Dekkera to produce volatile phenols has been well demonstrated in the literature (Heresztyn 1986, Chatonnet et al. 1995, Edlin et al. 1995), this genus (genera) being considered the main organisms responsible. Other yeasts and bacteria have, however, been shown to metabolize hydroxycinnamic acids (Cavin et al. 1993, Chatonnet et al. 1993, 1995, Shinohara et al. 2000). Lactic acid bacteria are part of the natural flora of wines. Among the various genera within this group, Lactobacillus, Pediococcus, Oenococcus, and Leuconostoc are of major importance to winemaking (Davis et al. 1988, Dicks et al. 1995). Oenococcus oeni is the main species responsible for malolactic fermentation, which is generally considered beneficial to the quality of red wines and of some white wines (van Vuuren and Dicks 1993, Liu 2002). In addition to malolactic fermentation, the growth and metabolism of certain lactic acid bacteria can induce a range of compositional changes (such as acidification, ropiness, mousy taint, geranium off-flavor, and bitterness) that may negatively affect the quality of the final product (Sponholz 1993, Lonvaud-Funel 1999). Concerning the production of volatile phenols, Cavin et al. (1993) have shown that p-coumaric and ferulic acids were decarboxylated by Lactobacillus brevis, Lactobacillus plantarum, and Pediococcus pentosaceus. Chatonnet et al. (1995) reported that some strains of L. brevis and P. pentosaceus were capable of decarboxylating p-coumaric acid to form 4-vinylphenol as actively as Saccharomyces cerevisiae. Oenococcus oeni synthesized small quantities of 4-vinylphenol. Only L. plantarum was capable of producing ethylphenols, but in low concentrations when compared to Brettanomyces. Thus, it can be seen from the literature that certain lactic acid bacteria strains decarboxylate phenolic acids into vinylphenols; the capacity to produce ethylphenols seems to be much rarer in this group of organisms.
Further research is needed on the role of wine microorganisms, other than Brettanomyces/Dekkera, and possible interactions between them in the production of volatile phenols. The main objective of this work was to evaluate the ability of lactic acid bacteria, mainly those species associated with wine, to produce volatile phenols (4-vinyl and/or 4-ethyl derivatives).
Materials and Methods
Bacterial strains.
The bacterial strains used in this study include wine isolates, isolated by Couto and Hogg (1994), from the Escola Superior de Biotecnologia, Portuguese Catholic University, Porto, culture collection, and commercial culture collection strains as indicated by their designations in Table 1⇓.
Production of 4-vinylphenol (4-VP) and 4-ethylphenol (4-EP) from p-coumaric acid, and 4-vinylguaiacol (4-VG) and 4-ethylguaiacol (4-EG) from ferulic acid by lactic acid bacteria, in MRS/tomato juice broth (50:50) (pH 4.5, 5% v/v ethanol).
Growth conditions and production of volatile phenols in synthetic medium.
The liquid growth medium (MRS/TJ) was a mixture (50:50) of two commercial media: MRS (de Man, Rogosa, and Sharpe) from Biokar Diagnostics (Beauvais, France) and TJ (tomato juice broth) from Difco (Detroit, MI). The pH was adjusted to 4.5 with a concentrated (6 M) hydrochloric acid solution before sterilizing. After sterilization (121°C, 15 min), ethanol (99.5% v/v) was added to the medium to obtain a final concentration of 5% v/v ethanol, which has been found to stimulate growth of wine lactic acid bacteria (Couto 1996).
Cultures were grown statically at 25°C to late exponential phase in MRS/TJ containing hydroxycinnamic acids at 500, 50, and 5 mg/L (3.05, 0.305, and 0.031 mM for p-coumaric acid; 2.57, 0.257, and 0.026 mM for ferulic acid). Experiments were also conducted with added 4-vinylphenol, at 100 mg/L, in the absence of phenolic acids. Each individual assay was performed in duplicate. p-Coumaric (p-hydroxycinnamic) acid, ferulic (p-hydroxymethoxycinnamic) acid, and 4-vinylphenol were obtained from Sigma-Aldrich (Steinheim, Germany). Fresh concentrated solutions (10 g/L) of phenolic acids and 4-vinyl-phenol were prepared in pure (99.5% v/v) ethanol and added to the growth media at the desired concentrations. Cultures were maintained on slants prepared with MRS/TJ with 2% agar (Lab M, Bury, UK), pH 4.5, at 4°C, with monthly transfers to maintain strain viability.
Analysis of volatile phenols.
Samples (50 mL) were taken at late exponential growth phase (5 to 7 days) from culture media and were centrifuged at 8000 x g for 10 min. Volatile phenols in the culture supernatants were analyzed according to the method developed by Bertrand (1981). A sample of 50.0 μL 3-octanol (internal standard) was added to 50 mL of culture medium. This mixture was successively extracted with 4.0 mL, 2.0 mL, and 2.0 mL of ether/hexane (1:1, v/v) by stirring for 5 min. The organic phases were collected, mixed, and concentrated under a stream of nitrogen to approximately one-third of the original volume. A sample of 2 μL of the extract was injected into a gas chromatography-flame ionization detector (Perkin Elmer, Shelton, CT) with a FFAP-type column (BP 21, 50 m x 0.25 mm x 0.2 μm) (SGE, Austin, TX). The injector (split/splitless) was heated to 250°C with a split flow of 30 mL/min and a splitless time of 0.3 min. The carrier gas flow was adjusted to 1 mL/min. The temperature of the oven was maintained at 40°C for 5 min after the injection and then increased at a rate of 2°C min−1 up to 220°C. This temperature was then maintained for 20 min. The standard error of the method is ~10 μg/L.
Results and Discussion
Production of volatile phenols in culture medium.
Representative species and strains of lactic acid bacteria associated with wine and other environments (Table 1⇑) were surveyed for the capacity to produce volatile phenols as evaluated by the gas chromatographic detection of p-coumaric and ferulic acid derivatives in synthetic culture medium. The phenolic composition of wine is complex and includes hydroxycinnamic acids in concentrations ranging from 100 to 200 mg L−1 (gallic acid included), depending on grape variety and vinification processes (Reguant et al. 2000). The culture conditions and the level of 500 mg/L of precursor compounds used for the lactic acid bacteria screening were chosen to facilitate bacterial growth and the olfactory detection of volatile phenols, respectively. 4-Vinylphenol and 4-ethylphenol, as products of p-coumaric acid metabolism, and 4-vinylguaiacol and 4-ethylguaiacol, as products of ferulic acid metabolism, were analyzed in cultures grown to late exponential phase. Experiments were performed in duplicate, but since results within each duplicate were similar (standard deviation never varied more than 10% of the mean value), data in Table 1⇑ refer to one assay. As shown in Table 1⇑, certain strains of lactic acid bacteria are able to produce volatile phenols. Thirteen strains (37%) produced volatile phenols from p-coumaric acid, although only three (9%) produced 4-ethylphenol, the final product of the p-coumaric acid metabolic pathway. The reduction step was only found in the Lactobacillus genus: L. brevis, L. collinoides, and L. plantarum (the two p-coumaric acid derivatives being found in the medium inoculated with L. plantarum NCFB1752). The other 10 strains were only capable of producing the intermediate compound, 4-vinylphenol. p-Coumaric and ferulic acid derivatives were not detected in media not supplemented with the phenolic acids and in media supplemented but not inoculated (data not shown).
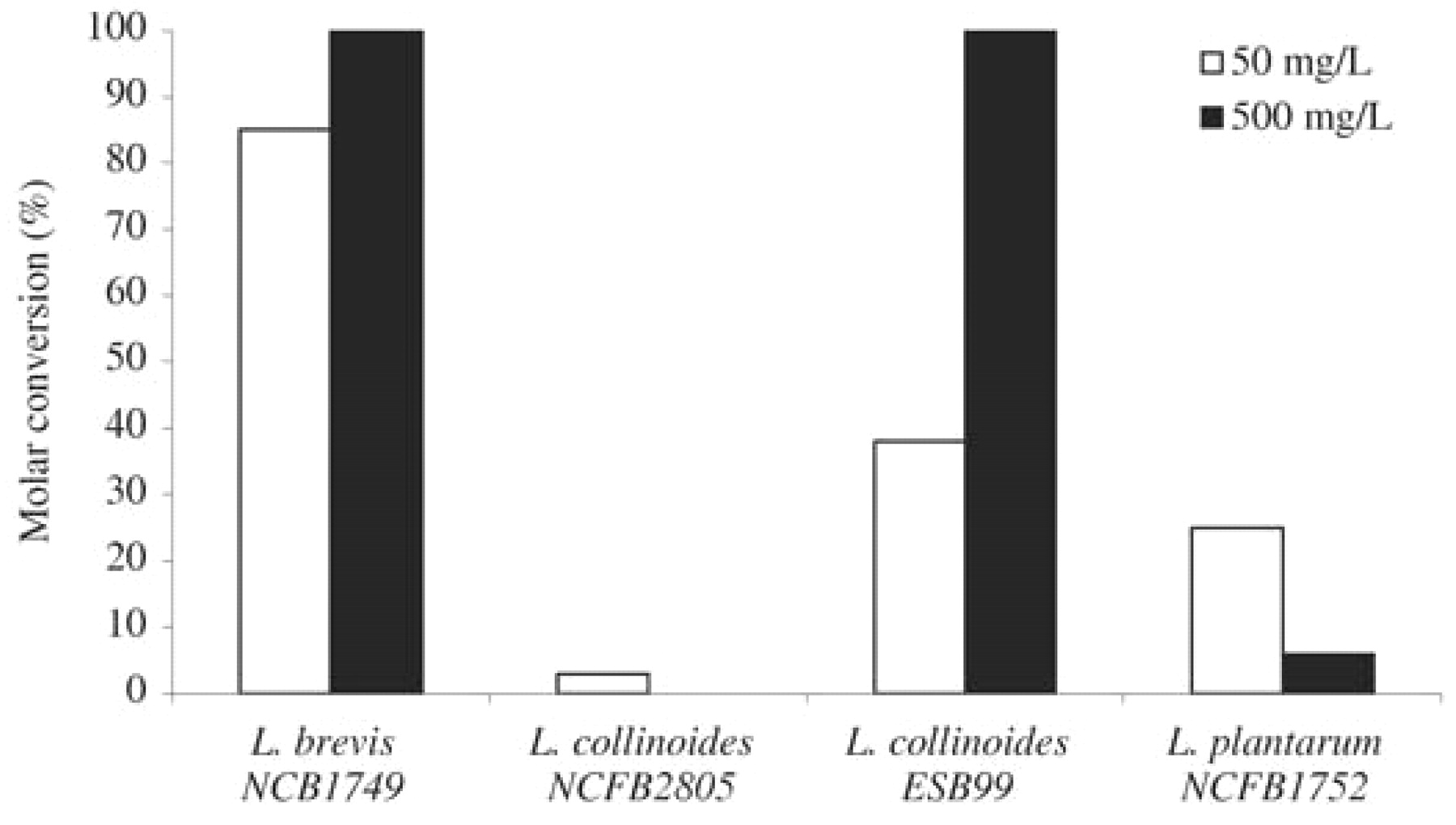
In agreement with the present results, Cavin et al. (1993) have reported that one strain of L. brevis and L. plantarum produced derivatives of p-coumaric and ferulic acids identified as 4-ethylguaiacol and 4-ethylphenol in culture medium. Lactobacillus plantarum was also reported to produce 4-ethylphenol (Chatonnet et al. 1995, Cavin et al. 1997, Barthelmebs et al. 2000a). However, Chatonnet et al. (1997) found that the level of volatile phenols produced by L. plantarum was significantly reduced when grown in wine in comparison to culture media. This reduction was associated with the presence of phenolic compounds, such as tannins, in wine that may exert an inhibitory action toward the cinnamate decarboxylase activity of this organism. Beek and Priest (2000) have shown that, similar to L. plantarum, L. crispatus possesses both decarboxylation and reduction activities. We show here that a strain of L. collinoides is a strong 4-ethylphenol producer, with a molar conversion rate of 100% in the conditions employed. Other lactobacilli species (L. mali, L. sake, and L. viridescens) produce 4-vinylphenol at high conversion rates, thus exhibiting decarboxylation activity but not reduction activity. The strains of L. hilgardii, the predominant ethanol tolerant species found in Port wine (Couto and Hogg 1994), assayed in this study did not show decarboxylation or reduction activity, which agrees with the results obtained for this species by Beek and Priest (2000).
Seven of the eight pediococci strains studied were able to produce 4-vinylphenol, but not 4-ethylphenol, from p-coumaric acid. Other researchers found the same type of activity in P. pentosaceus (Cavin et al. 1993, Chatonnet et al. 1995, Barthelmebs et al. 2000b). Our work shows that P. acidilactici and P. damnosus are also 4-vinylphenol producers, as also reported by Chatonnet et al. (1997) for the latter species. The impact of vinylphenols in wines has been frequently neglected, with more attention being paid to the influence of ethylphenols. It has been noted, however, that concentrations greater than 770 μg/L of a mixture of 4-vinylguaiacol/4-vinylphenol (1:1) in white wines can be responsible for phenolic and pharmaceutical off-odors (Chatonnet et al. 1993). The impact of the presence of vinylphenols in red wines should be further evaluated.
The two O. oeni strains studied did not produce either of the two p-coumaric acid derivatives. Decarboxylation activity in this species was found only when cells were submitted to permeabilization, thus indicating the lack of a transport system for the hydroxycinnamic acids (Cavin et al. 1993). The strain of Leuconostoc mesenteroides tested is also a nonvolatile phenols producer. Growth of this strain, of O. oeni, and of some of the lactobacilli strains studied seemed to be negatively affected by the presence of phenolic acids, however, as indicated by the relatively low final culture absorbance measured spectrophotometrically (Table 1⇑). Hydroxycinnamic acids at 500 mg/L may exert some inhibitory effect on lactic acid bacterial growth (Campos et al. 2003, Stead 1993).
The capacity of lactic acid bacteria to produce volatile phenols from ferulic acid was also assessed and was found to be much lower than from p-coumaric acid (Table 1⇑), which agrees with other findings (Chatonnet et al. 1995, Beek and Priest 2000). Most strains were able to convert ferulic acid to 4-vinylguaiacol and 4-ethylguaiacol, but at quite low conversion rates. The attained concentrations are, however, in some cases, above the perception threshold (130 μg/L for 4-vinylguaiacol and ~50 μg/L for 4-ethylguaiacol) (Chatonnet et al. 1992).
Influence of hydroxycinnamic acid concentration.
Strains found to be capable of producing volatile phenols were used in subsequent studies using lower phenolic acid concentrations. The higher the p-coumaric acid content in the culture medium, the more volatile phenols were produced (data not shown). We also observed that the molar conversion yield was dependent on the concentration of the substrate. With the production of 4-vinylphenol, higher conversion yield values were obtained with 500 mg/L of p-coumaric acid than with 50 mg/L (Figure 1⇓) or 5 mg/L (data not shown) in all strains studied. The decrease of the conversion yield with the substrate concentration appeared strain dependent. The same behavior was obtained in the production of 4-ethylphenol (Figure 2⇓), except for L. plantarum NCFB 1752, for which a higher conversion yield was obtained when cells were grown in the presence of 50 mg/L p-coumaric acid rather than 500 mg/L. The behavior of this strain, including the result obtained in the presence of 5 mg/L of p-coumaric acid, is shown in Figure 3⇓. The conversion yield of p-coumaric acid to 4-vinylphenol increased with the concentration of the substrate while the conversion to 4-ethylphenol decreased. It appeared that the production of 4-ethylphenol, in relation to 4-vinylphenol, was favored at low concentrations of p-coumaric acid.
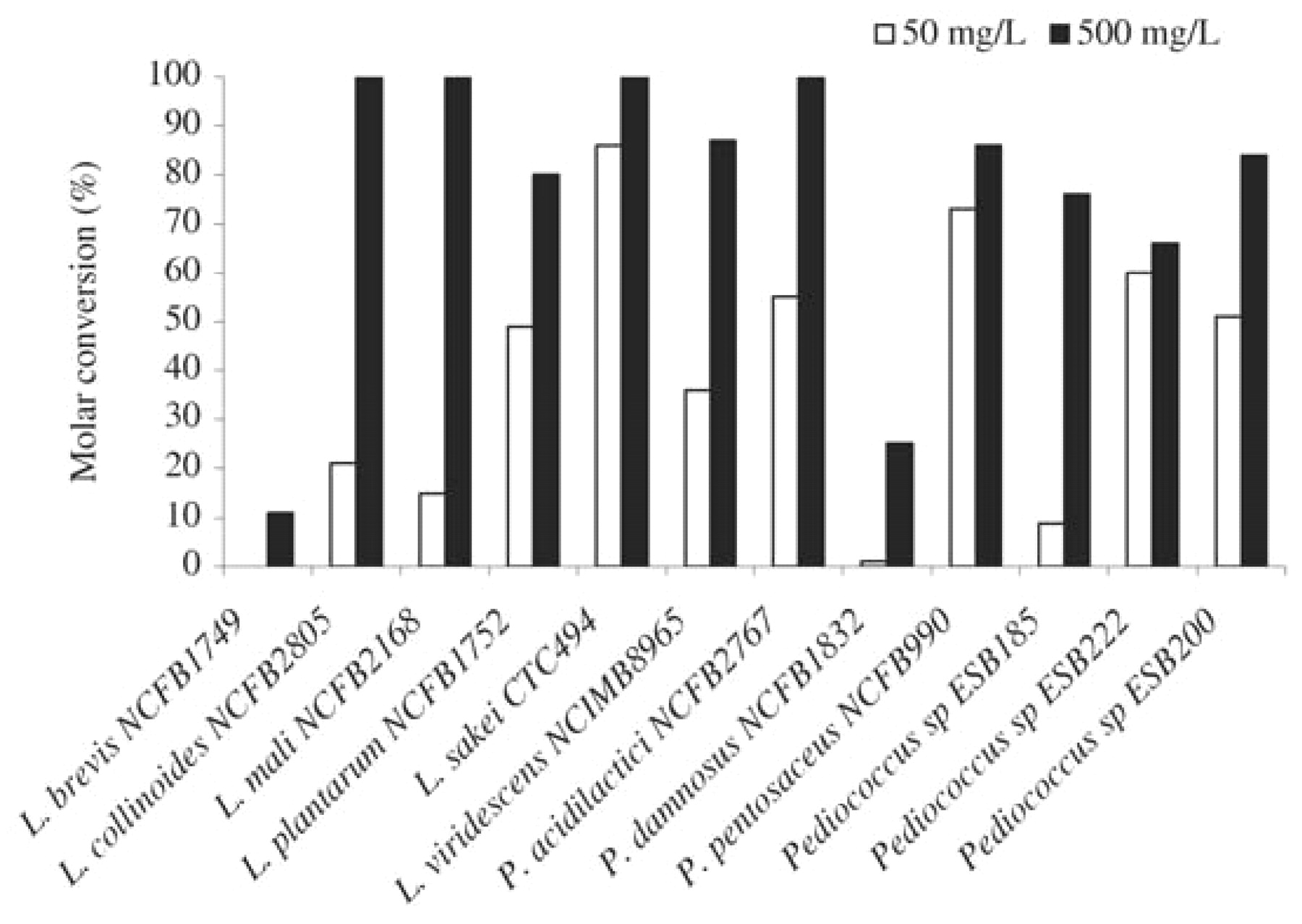
Molar conversion yield of p-coumaric acid (50 mg/L and 500 mg/L) into 4-vinylphenol.

Molar conversion yield of p-coumaric acid (50 mg/L and 500 mg/L) into 4-ethylphenol.

Molar conversion values of p-coumaric acid, at different concentrations, by L. plantarum NCFB1752 into 4-vinylphenol and to 4-ethylphenol.
The phenolic acid catabolism of L. plantarum is well documented in the scientific literature. An inducible p-coumaric acid decarboxylase (PDC) from this species was purified and characterized by Cavin et al. (1997), displaying a much higher activity on p-coumaric acid than on ferulic acid. The existence of another inducible acid phenol decarboxylase (PDC2), which has a lower activity than PDC, has been demonstrated (Barthelmebs et al. 2000a). In addition to these two enzymes, the existence of p-coumaric and ferulic acid reductase activities (PAR) has been revealed (Cavin et al. 1997, Barthelmebs et al. 2000a). These authors consider the PDC activity to be a stress response induced by the phenolic acid toxicity, consisting of the conversion of p-coumaric acid to 4-vinylphenol, a less toxic compound. Toxicity of phenolic acids is thought to be due to the dissipation of the cell ΔpH (uptake of the protonated form and the consequent release of H+ in the interior of the cell) and to a specific mechanism of phenolic acids. The PDC2 and PAR activities are less efficient as detoxification systems, their biological significance remaining unclear.
Influence of precursor compound on 4-ethylphenol.
As noted above, the production of volatile phenols is the outcome of the sequential activity of two enzymes (cinnamate decarboxylase and vinylphenol reductase). The intermediary metabolite, 4-vinylphenol, was tested as substrate for the production of 4-ethylphenol, in the absence of p-coumaric acid, in those strains that were not able to produce any of the volatile phenol compounds from p-coumaric acid. The strains studied were L. fructivorans ESB92; L. brevis LMG7934; L. hilgardii ESB5, ESB17, and ESB34; O.oeni VF and ESB132; P. parvulus LMG11486; and Leuc. mesenteroides NCFB523. None of these bacteria were able to form 4-ethylphenol from the metabolism of the intermediary compound, indicating that cinnamate decarboxylase and vinylphenol reductase are not present or active in these bacteria. The lack of a transport system for 4-vinylphenol can also be considered as a justification for the results obtained.
Conclusions
Results show that several species of lactic acid bacteria are capable of producing volatile phenols, particularly from the metabolism of p-coumaric and ferulic acids in culture medium. The survey performed in this work is, to our knowledge, the most extensive to date, employing 20 different lactic acid bacteria species (35 strains), most associated with wine or the wine environment. In the conditions used, significant quantities of volatile phenols were produced. Experiments with low added levels of p-coumaric acid (5 mg/L) to the culture medium showed that some strains can still produce relatively high concentrations of volatile phenols. In agreement with previous studies, most of the strains produced 4-vinylphenol and not 4-ethylphenol, thus exhibiting decarboxylation but not reduction activity in the metabolism of p-coumaric. Both activities, and the consequent formation of 4-ethylphenol, were detected only in lactobacillli strains. Lactobacillus collinoides is reported as volatile phenols producer, although strain specificity concerning this capacity was observed. Lactobacillus mali, L. sake, L. viridescens, and P. acidilactici were also found to be able to produce volatile phenols, although only capable of performing the decarboxylation step.
Among the main volatile phenols producers found in this study, the following species have been encountered in wine, thus being relevant to winemaking: L. brevis, L. collinoides, L. plantarum, L. mali, L. sake, P. damnosus, and P. pentosaceus. The capacity of these organisms to produce volatile phenols in wine or near wine conditions will be evaluated in future studies. The factors related to wine production and to the wine characteristics that may affect the enzymatic activities involved in the formation of volatile phenols are not yet well established. Moreover, more information is needed about the potential risk of the activity of these organisms, in terms of production of volatile phenols, during malolactic fermentation and in the postfermentation period.
Footnotes
Acknowledgments: The authors thank the Fundação para a Ciência e Tecnologia (FCT) for funding this research via projects POCTI/AGG/40724/2001 and POCI/AGR/61331/2004. F.M. Campos extends appreciation for the grant PRAXIS XXI/BD/19909/99.
Pedro Mota and Nuno Carvalho are thanked for helpful technical assistance.
- Received November 2005.
- Revision received February 2006.
- Copyright © 2006 by the American Society for Enology and Viticulture
Literature Cited
Vol 57 Issue 2








{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
More from this TOC section
Similar Articles
